Inledning
Kloroplaster utgör en strukturell funktion av växtceller och stöd växter överlevnad via deras primära ämnesomsättning och hög nivå funktioner (Kirk och Tilney-Bassett, 1978; Mulle, 1988; López-Juez och Pyke, 2005). Under växt vegetativ tillväxt innehåller bladceller en mycket homogen population av kloroplaster med avseende på storlek och form., Antalet kloroplaster per cell uppnås genom binär fission av befintliga organeller och uppdelning i två dottercellerna vid celldelningen (Birky, 1983; Possingham och Lawrence, 1983). Således är reglering av kloroplastnumret i en given celltyp avgörande för cellfunktionen och genetiskt arv av kloroplaster.
för att undersöka arten av kloroplastnummerbestämning i växtceller har bladmesofyllceller av representativa arter spelat en viktig roll ., Dessa celler är fysiologiskt viktiga för fotosyntes och visar en hög grad av strukturell och funktionell homogenitet. Tidig systematisk observation analyser av isolerade vävnader och celler (Boasson och Laetsch, 1969; Possingham och Saurer, 1969; Boffey et al., 1979; Lamppa m.fl.,, 1980; Thomas och Rose, 1983; Pyke och Leech, 1991) gav mycket användbar information om kloroplastnummerbestämning, inklusive uppfattningen att kloroplaster (plastider) inte syntetiseras de novo men replikeras genom division och observationen att leaf mesofyllkloroplastnummer är känsligt för olika miljö-och växt endogena faktorer. När det gäller den senare, i spenat, ljus har en positiv inverkan på kloroplast division under bladskivkultur jämfört med dim eller mörka förhållanden (Possingham och Lawrence, 1983)., I de första bladen av vete är cellvolymen positivt korrelerad med kloroplastproliferation (Ellis och Leech, 1985; Pyke och Leech, 1987). I Arabidopsis, den genetiska bakgrunden påverkar kloroplast spridning. den genomsnittliga kloroplast antal per cell i första bladen är 121 i Landsberg erecta (Ler) ecotype och 83 i Wassilewskija (Ws) ecotype (Pyke och Leech, 1994; Pyke et al., 1994)., Leaf mesofyllceller har också bidragit till att förstå den genetiska kontrollen av kloroplast division; till exempel, screening mutanter nedsatt i kloroplastproliferation och karakterisera genfunktioner involverade i kloroplast division har avslöjat över 20 gener som kodar för kloroplast division maskinkomponenter eller kloroplast reglerande faktorer (Gao och Gao, 2011; Miyagishima et al., 2011; Basak och Møller, 2013; Osteryoung och Pyke, 2014; Li et al., 2017).
däremot studier av replikation av kloroplaster i icke-mesofyllceller (t. ex.,, trottoarceller i blad epidermis; Itoh et al., 2018) är knappa. Nyligen har regleringen av kloroplast division rapporterats skilja sig mellan bladvävnader (Fujiwara et al., År 2018. Itoh et al., 2018), även om den detaljerade mekanismen fortfarande är okänd. Dessutom, medan analyserna av suspension-odlade Bi-2 celler och blad mesofyll protoplaster i tobak och skjuta apikala meristem och blad primordiala celler i Arabidopsis (Nebenführ et al., 2000; Sheahan m.fl.,, 2004; Seguí-Simarro och Staehelin, 2009) har gett viktiga insikter, hur kloroplast (plastid) partitionering är reglerad i växter är fortfarande oklart. Således, trots stora ansträngningar, grundläggande frågor i kloroplastforskning kvar, såsom (i) hur är kloroplast Antal per cell samordnas i växtvävnader och (ii) hur är kloroplast partitionering regleras vid celldelning.,
historia av forskning om Guard Cell Chloroplast Number
Stomatal GCs I shoot epidermis innehåller i allmänhet kloroplaster och kontroll gasutbyte mellan blad mesofyll och atmosfären (Sachs, 1875; Taiz et al. År 2015, se Figur 1A). Den första undersökningen av GC kloroplast nummer i blad utfördes över ett sekel sedan i naturligt odlade Drosera växter (Macfarlane, 1898)., Denna studie visade att, liksom andra växt-och cellkonstruktionsegenskaper, GC-kloroplastnummer per cell i en putativ hybrid härrörande från en korsning mellan Drosera filiformis och Drosera intermedia var mellanliggande mellan de två arterna, vilket innebär att GC-kloroplastnummer kunde användas för att bestämma en växts genetiska smink. Viktiga observationer rapporterades därefter om skillnaderna i GC-kloroplasttal bland växtarter (Sakisaka, 1929) och det relativt stabila kloroplasttalet i GCs i Mulberrys lövhud (Morus spp.,; Hamada och Baba, 1930) och i mogna blad av flera Brassica arter (Iura, 1934). Vidare visade analys av autopolyploida sockerbetor (Beta vulgaris) växter att GC-kloroplasttalet i löv är positivt korrelerat med kärnkraftsploidy-nivån av växter (Mochizuki och Sueoka, 1955). Mer djupgående och omfattande analyser genomfördes sedan med hjälp av olika växtprover för att undersöka förhållandet mellan kloroplastnummer och stomatalstorlek med ploidy-nivån (t.ex. Frandsen, 1968)., I dessa analyser antogs ofta kloroplasträkning vid stomatal (GC-par) – nivån, vilket utesluter effekten av partisk kloroplastfördelning mellan Parade GCs (t.ex. Mochizuki och Sueoka, 1955; Frandsen, 1968), vilket avslöjar att det genomsnittliga GC-kloroplasttalet i löv eller cotyledoner hos cirka 80 arter, varianter eller hybrider varierade från 2,8 till 40,0 i diploider (2×) och 5,0 till 73,5 i tetraploider (4×). Dessutom, hela genomet duplicering händelser i växter (dvs. 1× till 2×, 2× till 4×, etc.) orsakade en ungefär 1.,7-faldig ökning av GC-kloroplastnummer med hög trohet (granskad i Butterfas, 1973). Dessa resultat uppmuntras utredningar ploidy nivå i olika vävnader och växter som erhållits via vävnadsodling, korsning, eller naturlig odling, i kombination med kemiska (t ex, kolkicin) eller strålning behandlingar (t ex, Jacobs och Yoder, 1989; Singsit och Veilleux, 1991; Qin och Rotino, 1995)., Medan GC-kloroplastnummer har studerats i stomatalbiologi (Lawson, 2009) och cytologi för att förstå kloroplastmultiplikation (Butterfas, 1979; se nedan), har det i stor utsträckning fungerat som en pålitlig och bekväm markör för detektering av hybrider, arter och varianter och för uppskattning av ploidynivåer av målväxtvävnader.
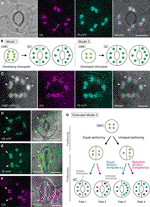
Figur 1 förenklade modeller och mikroskopibevis för kontroll av kloroplastnummer i stomatalskyddsceller (GCs)., (A) En typisk stomi (GC par) i abaxial epidermis hos Arabidopsis bladet uttrycka en stomi-riktade fusion av transitering peptid (TP) med cyan fluorescerande protein (GFP; TP-GFP). B) två modeller för bestämning av kloroplastnummer i GCs, som inbegriper antingen kloroplastfördelning (modell 1) eller både kloroplastproliferation och partitionering (modell 2) under GC-utveckling från guard mother cells (GMC). (C) EN GC par i adaxial epidermis av Arabidopsis blad bladskaft uttrycka TP-gemensamma FISKERIPOLITIKEN och FtsZ1 smält till grönt fluorescerande protein (GFP; FtsZ1-GFP)., (D–F) GC par i abaxial epidermis av Arabidopsis bladet med (D, E) eller utan (F) uttryck för TP smält den gula fluorescerande protein (YFP; TP-YFP). F) klorofyll-autofluorescens (Chl) användes som kloroplastmarkör., G) utökad modell 2, som representerar medverkan av lika och ojämn kloroplastfördelning efter GMC-uppdelning och efterföljande uppdelning av GC-kloroplaster med lika (blå linje) eller selektiv (röd linje) divisionskompetens, vilket skulle resultera i fyra typer av kloroplastnummesbestämning (Fates 1-4) under sen stomatal utveckling av Arabidopsisblad. (A, C-F) Epifluorescensmikroskopi utfördes med ett Olympus IX71 inverterat mikroskop med användning av växtmaterial som tidigare beskrivits (Fujiwara et al., 2017, Fujiwara et al., 2018)., Fluorescenssignaler av klorofyll, CFP, GFP och YFP är pseudofärgade i magenta, cyan, Grön (endast i sammanslagen bild) respektive grön. Indikationer i paneler är följande: pilhuvud, ftsz1-ringen; pil, förstorad GC kloroplast; streckad linje, cellform. Skala bar = 10 µm.
användbarhet av Bladskyddsceller för analys av Kloroplastnummerkontroll
Bladmesofyllceller har länge använts som en primär modell för analys av kloroplastnummer., Samtidigt som de har fördelar för den studie av effekterna av miljöförhållandena på kloroplast division (t ex, ljus-cytokinin signalering, Boasson och Laetsch, 1969; Possingham och Lawrence, 1983; Okazaki et al., 2009; Chiang et al., 2012), de är begränsade i vissa avseenden. För det första varierar bladmesofyllceller i storlek och form och fördelas djupt inom bladet, vilket gör det svårt att manipulera intakta vävnader., För det andra kan mottagligheten av bladmesofyllkloroplastproliferation för miljöbelastning och växttillväxtförhållanden förhindra tillförlitliga jämförelser mellan studier. Bladet mesophyll kloroplast antal per cell i Arabidopsis Columbia (Col) ecotype har rapporterats bland annat som 76 (Frände och Pyke, 1998), 80-100 (Stokes et al., 2000), 70 (Tirlapur och König, 2001), 41 (Yoder et al., 2007), 40-60 (Okazaki et al., 2009), och 30-40 (Kawade et al., 2013)., Vidare är det nästan omöjligt att bedöma kloroplastpartitioneringens bidrag till det slutliga kloroplasttalet per mesofyllcell under bladutvecklingen, även om detta tros bestämmas av balansen mellan celldelningshastigheten och kloroplastdivisionen. För att avslöja mekanismen för kloroplastnummerkontroll i vegetativa bladceller krävs ett modellsystem som övervinner ovanstående problem.
Stomatal GCs (se Figur 1A) uppvisar egenskaperna hos ett modellsystem för att förstå mekanismen för kloroplastnummerkontroll., GCs är mycket likformiga i storlek och form inom en vävnad, och deras spridda men täta fördelning i det yttersta skiktet av skott underlättar deras upptäckt genom ljus och fluorescensmikroskopi. GCs är också härstammar från protodermal celler i skjuta apikala meristem eller från embryonala epidermala celler, och deras utvecklings-sekvens genom meristemoids (en stomatala föregångare med meristematic aktivitet) och vakta mamma celler (GMCs, en föregångare till GC par) är etablerad i detalj (Zhao och Säck, 1999; Nadeau och Säck, 2002; Kalve ’ et al., 2014)., Sen stomatal utveckling innebär en enda runda av symmetrisk GMC-division, vilket möjliggör bedömning av kloroplastfördelning och partitionering före och efter cytokinesi. Ur praktiska experiment är blad GCs lämpliga för mikroskopi. Det visades tidigare att kloroplastnummer per cell i Bladgcs av Sinapis alba påverkades mindre av olika ljusförhållanden än i bladmesofyllceller (Wild and Wolf, 1980)., Dessutom är skillnaden i gc kloroplasttal i blad petioler relativt liten bland de tre Arabidopsis-ekotyperna Col, Ler och Ws (Fujiwara et al., 2018). Vidare har endoreduplication, som påverkar utvecklingen av bladmesofyll, trottoar och trichomceller, inte detekterats i Arabidopsis leaf GCs (Melaragno et al., 1993), vilket skulle säkerställa tolkningarna av kloroplastnummerdata på 2C-nivån av celler., Tillsammans tyder dessa rapporter på att blad GCs är potentiellt en utmärkt modell för systematisk analys av kloroplastnummer dynamik i en viss cell härstamning.
Arabidopsis Leaf Guard-celler som modell för att studera kontrollen av Kloroplastnummer
i historien om GC chloroplast research har kloroplasträkning vid stomatal (GC-par) – nivån tjänat en lika viktig roll för att bestämma kloroplastnumret som att räkna på den enskilda GC-nivån. Båda metoderna producerar samma genomsnittliga kloroplastnummer (Butterfas, 1973)., När variationen i kloroplastfördelning i parade GCs och dess underliggande mekanism är föremål för fokus är detaljerad information om kloroplaster på den enskilda cellnivån, dvs deras storlek, form och intracellulär lokalisering, väsentlig. Kloroplast (plastid) proliferation under GMC–GC-differentieringen undersöktes tidigare i flera växtarter (Butterfass, 1973, Butterfass 1979)., Dessa studier föreslog två modeller för att bestämma det slutliga kloroplastnumret i GCs i olika växtarter( Figur 1b): en (modell 1; sockerbetor) innebär endast kloroplastpartitionering vid GMC-division, och den andra innebär inte bara kloroplastpartitionering utan även kloroplastproliferation under GC-utveckling.
i en tid präglad av molekylär genetik, genomik, cell imaging, och andra tvärvetenskapliga analyser, det finns många möjligheter för ytterligare karakterisering av kloroplast partitioneringsmekanismen., Arabidopsis leaf GCs kan vara ett av de bästa modellsystemen för detta ändamål. Flera studier har undersökt GC kloroplast-nummer i bladen eller hjärtbladen av Arabidopsis (Hoffmann, 1968; Pyke och Leech, 1994; Pyke et al., 1994; Robertson m.fl., 1995; Keech m.fl., 2007; Chen et al., 2009; Yu et al., 2009; Higaki et al., 2012; Fujiwara et al., 2018). Dessa GCs uppvisar ett blygsamt antal kloroplaster, som sträcker sig från 3,5 till 5,5 i genomsnitt. Hittills har inga studier undersökt förändringarna i kloroplast (plastid) antal under stomatal utveckling., Mikroskopiska bevis från stomatalutvecklingsanalyser (t.ex. Zhao och Sack, 1999; Hachez et al., 2011) och våra preliminära observationer tyder på att GMC kan innehålla mindre antal utvecklande kloroplaster än GCs och att kloroplastproliferation kan inträffa under GC-differentiering. För att testa detta övervakades bildandet av kloroplast-divisionsmaskineriet i GCs med sonden FtsZ1 smält till det gröna fluorescerande proteinet (Ftsz1-GFP) (Fujiwara et al., 2008)., En transgen linje, samtidigt som uttrycker en transit-peptid (TP)-smält gemensamma FISKERIPOLITIKEN och FtsZ1-GFP-att visualisera stroma och FtsZ1 ring, respektive, blev undersökt av epifluorescence mikroskopi (Fujiwara et al., 2017). Expanding leaf petioles (femte blad av 4 veckor gamla plantor) användes. Som ett resultat detekterades GCs med symmetriskt förträngande kloroplaster (figur 1C). Dessa kloroplaster bildade ftsz1 ringen, en kloroplast division ring på stromal ytan av det inre kuvertmembranet i blad mesofyll och trottoarceller (Vitha et al., 2001; Fujiwara m.fl.,, 2008), vid ekvatorialkonstriktionsplatsen. I överensstämmelse med stomatalmönstringen i Arabidopsis leaf development (Donnelly et al. 1999; Andriankaja m.fl., 2012), dividera kloroplaster upptäcktes i slutet, liksom tidigt, skede av blad expansion. Således är Modell 2 sannolikt den bästa passformen för Arabidopsis leaf GCs.
förhållandet mellan Kloroplastproliferation och Expansion I Arabidopsis Leaf Guard celler
vidare observerades en oväntad fenotyp av GC kloroplast morfogenes hos mogna GCs (figurerna 1D, E)., När epidermala skalar av fullt expanderade löv (tredje fjärde bladblad av 4 veckor gamla plantor) från en TP-smält gul fluorescerande protein (YFP) linje mikroskopiskt karakteriserades (FL6-5 linje; Fujiwara et al., 2018), vissa stomata visade ojämlika kloroplastfördelningsmönster i GC-par, medan de flesta blad stomata visade lika eller liknande kloroplastfördelningsmönster (Robertson et al., 1995; Fujiwara m.fl., 2018)., Inom GC-paret i en stomi var storleken på kloroplaster i GC som innehöll mindre antal kloroplaster större än i de andra GCs i paret som innehöll större antal kloroplaster (figurerna 1D, E). På detta sätt upprätthåller GCs förmodligen den totala kloroplastvolymen per cell på en konstant nivå under celltillväxt. Förstorade kloroplaster representerade den terminala fenotypen och kunde inte längre dela i expanderade löv. Dessa resultat bekräftades i flera oberoende experiment, oberoende av uttrycket av ett TP-smält fluorescerande protein för stroma-märkning (figur 1F).,
Denna GC kloroplast fenotyp tolkas som en kompensationsmekanism för kloroplast expansion, som var väl dokumenterade i löv mesophyll defekta celler i kontrollen av kloroplast division (Pyke och Leech, 1994; Pyke et al., 1994). Hittills har endast en studie (Ellis och Leech, 1985) rapporterat en negativ korrelation mellan kloroplastnummer och kloroplaststorlek i bladmesofyllceller av vete, medan många studier har rapporterat en positiv korrelation mellan cellvolym och kloroplastnummer i normala bladmesofyllceller (Leech och Pyke, 1988; Pyke, 1997)., Obalanser i GC-kloroplasttal uppträder vid låg frekvens (Fujiwara et al., 2018), chloroplast heterogenity i GC-par indikerar att ojämn kloroplastpartitionering kan utlösa differentiell kloroplasttillväxt mellan vilda bladceller i Arabidopsis, trots symmetrisk celldelning.
kloroplastkompensationseffekten i GCs kan vara mindre strikt än i bladmesofyllceller., GCs skulle kunna klara av en brist eller total förlust av den totala kloroplast volym per cell i kraftigt nedsatt kloroplast division mutanter, som i Arabidopsis arc6 och atminE1 och tomat suffulta, medan många mutant GCs visade på en minskad kloroplast-nummer och utvidgade kloroplast storlek på samma sätt som löv mesophyll-celler (Robertson et al., 1995; Ut och Pyke, 2006; Chen et al., 2009; Fujiwara et al., 2018)., I en sen kloroplastdivisionsmutant, arc5, var minskningen av GC-kloroplasttalet inte associerad med en signifikant ökning av kloroplaststorleken, till skillnad från i bladmesofyllceller (Pyke och Leech, 1994). En lägre grad av kloroplastexpansion i GCs än i mesofyllceller (Pyke och Leech, 1994; Barton et al., 2016), och variationen i kloroplastexpansion bland GCs, kan ligga till grund för ett så brett tillåtet intervall av total kloroplastvolym per GC., Dessutom kan tidpunkten för kloroplastdelning under GMC-GC-differentiering väsentligt påverka den terminala GC-kloroplastfenotypen. Även om ytterligare detaljerad karakterisering krävs för att ta itu med denna fråga, verkar det rimligt att Arabidopsis leaf GCs representerar ett system för att undersöka de outnyttjade aspekterna av kloroplastnummerkontroll i växtceller.,
en arbetsmodell för bestämning av Kloroplastnummer i Arabidopsis Leaf Guard-celler
på grundval av ovanstående föreslår vi en arbetsmodell (en utökad modell 2) för analys av kloroplastnummer i GCs (figur 1g). Det slutliga kloroplasttalet per GC bestäms genom kloroplastpartitionering vid GMC-division och kloroplastproliferation i GCs. Under GMC-divisionen kan kloroplaster genomgå antingen lika eller ojämn partitionering. Under kloroplastproliferation kommer GC-kloroplaster att föröka sig med antingen lika (blå linje) eller selektiv (magenta line) divisionskompetens., Till exempel, om lika partitionerade kloroplaster har motsvarande divisionskompetens, kommer lika kloroplasttal att inträffa i GC-paret (Fate 1). Om ojämnt fördelade kloroplaster har likvärdig divisionskompetens kommer kloroplaster att öka i samma takt inom GC-paret (Fate 3). Om selektiv kloroplastdelning sker i GCs, kommer balansen av kloroplastnummer i GC-paret att förändras efter GMC-division (Fates 2 och 4)., Det är för närvarande svårt att hitta stöd för ”selektiv kloroplastdelning”, men om Fates 1 och 4 faktiskt dominerar i GCs, kan de ha en mekanism som kontrollerar total kloroplastvolym per cell, som i bladmesofyllceller. Modellen ger upphov till två frågor: (i) är GC kloroplaster ordentligt partitionerade i dotterceller och hur partitionerar de? Och ii) är Divisionens kompetens hos GC-kloroplaster samordnat reglerad?,
När det gäller i), om kloroplast arv sker genom slumpmässig fördelning av flera kloroplaster i cytoplasman eller genom positiv kloroplastpartitioneringsmekanism(er) har varit en långvarig oro (Butterfass, 1969; Birky, 1983; Hennis och Birky, 1984; Nebenführ, 2007; Sheahan et al., 2016). Intriguingly, i Arabidopsis arc6, blad eller cotyledon GCs har noll till tre kloroplaster, och i kloroplast-bristfälliga GCs, icke-fotosyntetiska plastider fortfarande finns i vesikulär till långsträckta former (Robertson et al., 1995, Chen m.fl., 2009; Fujiwara et al., 2018)., Inga GCs som saknar plastider i sig har hittats i arc6, och ingen förklaring till detta har kommit, trots störningen av kloroplastdivisionsapparaten (Vitha et al., 2003). Följaktligen kommer det att vara viktigt att undersöka replikation och morfologi av arc6 kloroplaster i stomatal härstamning studier. Arabidopsis mutant forskning kan också ge en annan ledtråd för denna fråga., Observationen att 18% av hjärtblad GCs i den skrynkliga blad (crl) mutant innehåller inga plastidic strukturer i cytoplasman, medan 100% av löv mesophyll celler innehålla en till fyra utvidgade kloroplaster (Asano et al., 2004; Chen et al., 2009), är av stor betydelse. CRL är ett kloroplastprotein med okänd funktion. Förstå CRL protein funktion kan ge insikter i mekanismen(er) av kloroplast partitionering., Analysen av kloroplastproliferation och partitionering i bladmesofyllceller i Arabidopsisbågsmutanter och andra transgena linjer har främjat forskning om proliferation och partitionering av icke-mesofyllplastider. På samma sätt kan resultat som erhållits i GCs överföras till andra cellsystem.
slutlig anmärkning
GC-modellen öppnar många möjligheter till utveckling av kloroplastbiologi. Till exempel, medan cytoskeletala system är kända för att reglera kloroplast morfologi, rörelse och partitionering (Sheahan et al.,, 2016; Wada, 2016; Erickson och Schattat, 2018), rollen för varje regulatorisk gen i kloroplastproliferation och partitionering i växter har fått liten uppmärksamhet. Å andra sidan, när det blir möjligt att försämra GC kloroplast nummer eller morfologi via olika experimentella strategier, nya insikter i molekylär kontroll av kloroplast morfogenes i stomatal härstamning celler kan tillhandahållas. Dessutom, i samband med kvantitativa analyser av kloroplastnummer under stomatal utveckling, kan matematisk modellering erbjuda en ny väg för dessa undersökningar., Detta dokument presenterar aktuell kunskap om hur GC kloroplast nummer styrs och belyser den potentiella nyttan av Arabidopsis leaf GCs för att förstå kloroplastproliferation och partitionering.
datatillgänglighet uttalande
alla datauppsättningar för denna studie ingår i artikeln / kompletterande material.
författare bidrag
MF tänkt studien och skrev manuskriptet. MF och som genomfört experimenten. AS och RI genomförde analyserna. Alla författare läste och godkände det slutliga manuskriptet.,
Finansieringen
Detta arbete fick stöd av Ministeriet för Utbildning, Kultur, Vetenskap och Teknik i Japan under KAKENHI (bidrag nr. 19K05831 till MF och 18K06314 till RI).
intressekonflikt
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
bekräftelser
författarna tackar Dr.Nobuyuki Kanzawa (Sophia University) för råd om arbetet.
Birky, C. W., Jr (1983)., Partitioneringen av cytoplasmiska organeller vid celldelning. Int. Rev. Cytol. 15, 49-86.
Google Scholar
butter churn, T. (1969). Plastidfördelningen i mitos av stängningscellens moderceller av haploid swedenklee (Trifolium hybridum L.). Planta 84, 230-234. doi: 10.1007/BF00388108
PubMed Abstrakt | CrossRef Full Text | Google Scholar
smör churn, T. (1973). Kontroll av plastid division med hjälp av nukleär DNA-mängd. Protoplasma 76, 167-195. doi: 10.,1007/BF01280696
CrossRef Full Text | Google Scholar
Qin, X., Rotino, G. L. (1995). Chloroplast number in guard cells as ploidy indicator of in vitro-grown androgenic pepper plantlets. Plant Cell Tissue Organ Cult. 41, 145–149. doi: 10.1007/BF00051583
CrossRef Full Text | Google Scholar
Sachs, J. (1875). Textbook of Botany. Oxford: Clarendon Press.
Google Scholar
Thomas, M. R., Rose, R. J. (1983)., Plastid antal och plastid strukturella förändringar i samband med tobak mesofyll protoplast kultur och växtregenerering. Planta 158, 329-338. doi: 10.1007/BF00397335
PubMed Abstrakt | CrossRef Full Text | Google Scholar
Tirlapur, U. K., König, K. (2001). Femtosecond nära infraröda lasrar som ett nytt verktyg för icke-invasiv realtid högupplösta time-lapse avbildning av kloroplast division I levande bunt mantelceller av Arabidopsis. Planta 214, 1-10. doi: 10.,1007/s004250100597
PubMed Abstrakt | CrossRef Full Text | Google Scholar
CrossRef Full Text | Google Scholar
















Lämna ett svar