introduktion
den neurala bearbetningen av ansikten är ett allmänt undersökt ämne för kognitiv vetenskap. Baserat på funktionella avbildningsstudier, enstaka cellinspelningar och neuropsykologisk forskning har det föreslagits att Ansiktsbehandling utförs av ett distribuerat nätverk, som involverar flera kortikala områden i däggdjurshjärnan (Haxby et al., 2000; Marotta m.fl., 2001; Rossion et al., 2003a; Avidan et al., 2005; Sorger et al., 2007)., Medan omfattningen av detta ansiktsbehandlingsnätverk för närvarande är under intensiv debatt (Ishai, 2008; Wiggett och Downing, 2008) är de flesta forskare överens om att det finns många kortikala områden som aktiveras av ansikts stimuli. Den mest inflytelserika modellen av ansiktsuppfattning, baserad på den ursprungliga modellen av Bruce och Young, 1986; Young och Bruce, 2011) föreslår en skillnad mellan representation av invariant och variant aspekter av ansiktsuppfattning på ett relativt oberoende sätt, separerade i en ”kärna” och en ”utökad” del (Haxby et al., 2000)., De viktigaste regionerna i ”stomnätet” är områden i occipital och lateral fusiform gyri (FG). Områdena i dessa två anatomiska regioner verkar vara specialiserade för olika uppgifter: medan occipital ansiktsområdet (OFA), som ligger på den underlägsna occipital gyrus (IOG) verkar vara involverad i strukturell bearbetning av ansikten, verkar fusiform face area (FFA)-processerna på ett högre nivå sätt, vilket till exempel bidrar till behandlingen av identitet (Sergent et al., 1992; George m.fl., 1999; Ishai et al., 1999; Hoffman och Haxby, 2000; Rossion et al., 2003a,b; Rotshtein et al.,, 2005). Dessutom de föränderliga aspekter av ansikten (såsom ansiktsuttryck, riktning ögon-blick, uttryck, läpp rörelser (Perrett et al., 1985, 1990), eller läppläsning (Campbell, 2011) verkar behandlas i superior temporal sulcus (STS; Puce et al., 1998; Hoffman och Haxby, 2000; Winston et al., 2004). De tre ovan nämnda områdena (FFA, OFA, STS) utgör den så kallade ”kärnan” i det perceptuella systemet för ansiktsbehandling (Haxby et al., 2000; Ishai et al., 2005)., Medan grundläggande information om ansikten behandlas av detta kärnsystem komplex information om andras humör, intressenivå, attraktivitet eller uppmärksamhetsriktning lägger också till information för att möta uppfattningen och behandlas av ett extra, så kallat ”utökat” system (Haxby et al., 2000). Detta system innehåller hjärnregioner med ett stort antal kognitiva funktioner relaterade till behandlingen av utbytbara ansiktsaspekter (Haxby et al., 2000; Ishai et al., 2005) och inkluderar områden såsom amygdala, insula, sämre frontal gyrus liksom orbitofrontal cortex (Haxby et al.,, 2000, 2002; Fairhall och Ishai, 2007; Ishai, 2008).
interaktioner av de ovan nämnda områdena modelleras i den nuvarande studien med hjälp av metoder som beräknar den effektiva anslutningen bland kortikala områden. Dynamic causal modeling (DCM) är en allmänt använd metod för att utforska effektiv anslutning bland hjärnregioner. Det är ett generiskt tillvägagångssätt för modellering av det ömsesidiga inflytandet av olika hjärnområden på varandra, baserat på fMRI-aktivitet (Friston et al., 2003; Stephan m.fl., 2010) och för att uppskatta inter-anslutningsmönstret för kortikala områden., DCMs är generativa modeller av neurala reaktioner, vilket ger ”a posteriori” uppskattningar av synaptiska kopplingar mellan nervceller populationer (Friston et al., 2003, 2007). Förekomsten av det distribuerade nätverket för ansiktsbehandling bekräftades först genom funktionell anslutningsanalys av Ishai (2008), som hävdade att den centrala noden för ansiktsbehandling är lateral FG, ansluten till lägre orderområden i IOG samt till STS, amygdala och frontala områden. Det första försöket att avslöja ansiktsbehandlingsnätet vid realistiska dynamiska ansiktsuttryck gjordes av Foley et al. (2012)., De bekräftade IOG -, STS-och FG-områdenas roll och fann att anslutningsstyrkan mellan medlemmarna i stomnätet (OFA och STS) och det utökade systemet (amygdala) ökar för bearbetning av påverkande gester. För närvarande utarbetade flera studier vår förståelse på ansiktsbehandlingsnätet och avslöjade en direkt koppling mellan amygdala och FFA och rollen som denna koppling i uppfattningen av rädda ansikten (Morris et al., 1996; Marco m.fl., 2006; Herrington m.fl., 2011)., Slutligen var effekten av högre kognitiva funktioner på ansiktsuppfattning också modellerad genom att testa anslutningarna av orbitofrontal cortex till kärnnätet (Li et al., 2010). Den hittades (Li et al., 2010) att orbitofrontal cortex har en effekt på OFA, vilket ytterligare modulerar informationsbehandlingen av FFA.
medan tidigare effektiva anslutningsstudier avslöjade detaljerna i ansiktsbehandlingsnätet relaterat till olika aspekter av ansiktsuppfattning, ignorerade de det enkla faktum att ansikten också kan betraktas som visuella objekt., Vi vet från en stor mängd experiment att visuella objekt behandlas av ett distribuerat kortikalt nätverk, inklusive tidiga visuella områden, occipito-temporala och ventrala temporala kortikaler, som i stor utsträckning överlappar ansiktsbehandlingsnätet (Haxby et al., 1999, 2000, 2002; Kourtzi och Kanwisher, 2001; Ishai et al., 2005; Gobbini och Haxby, 2006, 2007; Haxby, 2006; Ishai, 2008). Ett av de viktigaste områdena för visuell objektbehandling är den laterala occipitala cortexen (LOC), som kan delas in i två delar: den främre ventrala (PF/LOa) och den caudal–dorsala delen (LO; Grill–Spector et al.,, 1999; Halgren m.fl., 1999). LOC beskrevs först av Malach et al. (1995), som mätte ökad aktivitet för objekt, inklusive kända ansikten också, jämfört med kodade objekt (Malach et al., 1995; Grillspector m.fl., 1998a). Sedan dess betraktas lateral occipital (LO) främst som ett objektselektivt område, vilket ändå alltid visat sig ha förhöjd aktivering för ansikten också (Malach et al., 1995; Puce m.fl. 1995; Lerner m.fl., 2001), särskilt för inverterade sådana (Aguirre et al., 1999; Haxby et al. Epstein m.fl.,, 2005; Yovel och Kanwisher, 2005).
Således är det ganska förvånande att medan flera studier har behandlat effektiv anslutning av ansiktsbehandlingsområden, ansåg ingen av dem att LO: s roll i nätverket var. I en tidigare fMRI studie fann vi att LO har en avgörande roll i sensorisk konkurrens för ansikts stimuli (Nagy et al., 2011)., Aktiviteten hos LO reducerades genom presentationen av samtidigt presenterade samtidiga stimuli och denna responsreduktion, som återspeglar sensorisk konkurrens bland stimuli, var större när den omgivande stimulansen var ett ansikte jämfört med en Fourierfas randomiserad brusbild. Detta resultat stödde också tanken att LO kan spela en särskild roll i ansiktsuppfattning. Därför undersökte vi uttryckligen, med hjälp av metoder för effektiv anslutning, hur LO är kopplad till FFA och OFA, medlemmar i det föreslagna stomnätet för ansiktsuppfattning (Haxby et al., 1999, 2000, 2002; Ishai et al., 2005).,
material och metoder
försökspersoner
tjugofem friska deltagare deltog i experimentet (11 kvinnor, median: 23 år, min. 19 år, max. 35 år). Alla hade normal eller korrigerad till normal syn (självrapporterad), ingen av dem hade några neurologiska eller psykiska sjukdomar. Ämnen gav sitt skriftliga informerade samtycke i enlighet med de protokoll som godkänts av den etiska kommittén vid universitetet i Regensburg.,
Stimuli
ämnen presenterades centralt av gråskaliga ansikten, icke-sense-objekt och de Fourier randomiserade versionerna av dessa stimuli, skapade av en algoritm (Nasanen, 1999) som ersätter fasspektrumet med slumpmässiga värden (från 0° till 360°), vilket lämnar amplitudspektrumet för bilden intakt, samtidigt som någon forminformation tas bort. Ansikten var full front digitala bilder av 20 unga män och 20 unga kvinnor. De passade sig bakom en rund formmask (3,5° diameter) som eliminerade ansiktets yttre konturer (se en provbild i Figur 1)., Objekten var oansenliga, renderade objekt (n = 40) med samma medelstorlek som ansiktsmasken. Luminans och kontrast (dvs standardavvikelsen för luminansfördelningen) hos stimuli likställdes genom att matcha luminanshistogrammen (medel luminans: 18 cd/m2) med hjälp av Photoshop. Stimuli projicerades tillbaka via en LCD-videoprojektor (JVC, DLA-G20, Yokohama, Japan, 72 Hz, 800 × 600 upplösning) på en genomskinlig cirkulär skärm (app. 30 ° diameter), placerad inuti skannerborren vid 63 cm från observatören., Stimulanspresentation kontrollerades via e-prime software (Psychological Software Tools, Pittsburgh, PA, USA). Ansikten, föremål och Fourierbrusbilder presenterades i efterföljande block av 20 s, interfolierade med 20 S tomma perioder (enhetlig grå bakgrund med en luminans på 18 cd/m2). Stimuli presenterades för 300 ms och följdes av ett ISI 200 ms (2 Hz) i slumpmässig ordning. Varje block upprepades fem gånger. Deltagarna ombads att kontinuerligt fokusera på ett centralt presenterat fixeringsmärke., Dessa funktionella localizer-körningar var en del av två andra experiment av ansiktsuppfattning, publicerad någon annanstans (Nagy et al., 2009, 2011).

Figur 1. Prov stimuli av experimentet. Alla bilder var gråskala, samma i storlek, luminans och kontrast. Vänster panel visar ett ansikte, könsspecifika funktioner (som hår, smycken etc.) gömdes bakom en oval mask. Mittenpanelen visar ett prov icke-sense geometriskt objekt, medan höger panel visar Fourier-fas randomiserad version av objekt, som används som styrstimuli.,
datainsamling och analys
bildbehandling utfördes med hjälp av en 3-T MR Head scanner (Siemens Allegra, Erlangen, Tyskland). För den funktionella serien vi kontinuerligt förvärvade bilder (29 skivor, 10° lutas i förhållande till axiell, T2 * viktade epi sekvens, TR = 2000 ms; TE = 30 ms; flip vinkel = 90°; 64 × 64 matriser; i-plan upplösning: 3 mm × 3 mm; skiva tjocklek: 3 mm). Högupplösta sagittal T1-viktade bilder förvärvades med hjälp av en magnetiseringsepi-sekvens (MP-RAGE; TR = 2250 ms; TE = 2.,6 ms; 1 mm isotropisk voxel storlek) för att erhålla en 3D strukturell scan (för mer information, se Nagy et al., 2011).
funktionella bilder korrigerades för förvärvsfördröjning, omgjorda, normaliserades till MNI-utrymmet, omprofilerades till 2 mm × 2 mm × 2 mm Upplösning och rumsligt jämnas med en Gaussisk kärna av 8 mm FWHM (SPM8, Welcome Department of Imaging Neuroscience, London, UK; för detaljer om dataanalys, se Nagy et al., 2011).
VOI Selection
först valdes intressevolymer (VOI), baserat på aktivitet och anatomiska begränsningar (inklusive maskering för relevanta hjärnregioner)., Ansikts selektiva områden definierades som ett område som visar större aktivering för ansikten jämfört med Fourier brus bilder och objekt. FFA har definierats inom den Laterala Fusiform Gyrus, medan OFA inom IOG. LO definierades från objektet > Fourier brus och ansiktsbilder kontrast, inom mitten Occipital Gyrus. VOI-valet baserades på de t-kontraster som justerades med f-contrast (p < 0,005 okorrigerade med en minsta klusterstorlek på 15 voxels)., VOIs var sfäriska med en radie på 4 mm runt toppaktiveringen (för enskilda koordinater, Se tabell A1 i bilagan). Variansen som förklaras av den första egenvariaten av de djärva signalerna var över 79%. Endast högra halvklotområden användes i den nuvarande DCM-analysen, eftersom flera studier pekar på den dominerande rollen hos denna halvklot i ansiktsuppfattning (Michel et al., 1989; Sergent et al., 1992).,
en Effektiv Anslutning Analys
en Effektiv anslutning testades av DCM-10, som genomförts i SPM8 toolbox (Wellcome Institutionen för Bildbehandling Neurovetenskap, London, UK), som körs i Matlab R2008a (The MathWorks, Natick, MA, USA). Modeller av DCM definieras med endogena anslutningar, som representerar koppling mellan hjärnregioner (matris a), modulatoriska anslutningar (matris B) och drivingång (matris C). Här i A-matrisen definierade vi anslutningarna mellan de ansikts selektiva regionerna (FFA och OFA) och LO., Bilder av ansikten och objekt fungerade som driving input (matrix C) och vid detta analyssteg tillämpade vi inte några modulatoriska effekter på anslutningarna.
Modelluppskattning syftade till att maximera de negativa frienergiuppskattningarna av modellerna (F) för en given datauppsättning (Friston et al., 2003). Denna metod säkerställer att modellen passar använder parametrarna på ett parsimonious sätt (Ewbank et al., 2011). De uppskattade modellerna jämfördes, baserat på modellbevis p(y|m), vilket är sannolikheten p för att erhålla observerade data y som ges av en viss modell m (Friston et al., 2003; Stephan m.fl., 2009)., I denna studie tillämpar vi den negativa frienergi approximationen (variationsfri energi) till loggbeviset (MacKay, 2003; Friston et al., 2007). Bayesiansk Modell Urval (BMS) har utförts på både slumpmässiga (RFX) och fast (FFX) effekt mönster (Stephan et al., 2009). BMS RFX är mer motståndskraftig mot avvikare än FFX och det förutsätter inte att samma modell skulle förklara funktionen för varje deltagare (Stephan et al., 2009). Med andra ord är RFX mindre känslig för ljud., I RFX-tillvägagångssättet är analysens utgång den överskridande sannolikheten för modellutrymmet, vilket är i vilken utsträckning en modell är mer sannolikt att förklara de uppmätta data än andra modeller. Den andra produktionen av RFX-analysen är den förväntade bakre sannolikheten, vilket återspeglar sannolikheten för att en modell genererade de observerade uppgifterna, vilket möjliggör olika distributioner för olika modeller. Båda dessa parametervärden reduceras genom att modellutrymmet breddas (dvs.,, genom att öka antalet modeller), därför de beter sig på ett relativt sätt och modeller med delade funktioner och osannolika modeller kan snedvrida produktionen av analysen. Därför, förutom den direkta jämförelsen av de 28 skapade modellerna delade vi modellutrymmet i familjer, med liknande anslutningsmönster, med hjälp av metoderna för Penny et al. (2010).
Eftersom flera tidigare DCM studier pekar på att en nära dubbelriktad förbindelse mellan FFA och OFA (Ishai, 2008; Gschwind et al.,, 2012) i vår analys var dessa två områden alltid kopplade till varandra och LO var kopplad till dem på alla biologiskt rimliga sätt. De 28 relevanta modellerna delades in i tre modellfamiljer baserade på strukturella skillnader (Penny et al., 2010; Ewbank et al., 2011). Familj 1 innehåller modeller med linjära anslutningar mellan de tre områdena, förutsatt att informationen strömmar från LO till FFA via OFA. Familj 2 innehåller modeller med en triangulär struktur där LO skickar input direkt till FFA och OFA är också direkt kopplad till FFA., Familj 3 innehåller modeller, där de tre områdena är sammanlänkade och antar ett cirkulärt informationsflöde (Figur 2). För att begränsa antalet modeller i detta steg i analysen modulerade insatsvärdena enbart aktiviteten i deras införselområden. De tre familjerna jämfördes med en slumpmässig design BMS.
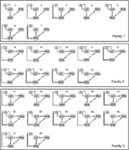
Figur 2. De 28 analyserade modellerna. Svarta linjer markerar uppdelningen mellan de tre familjerna, med olika matrisstrukturer. För mer information, se avsnittet ” material och metoder.,”
För det andra utarbetades modellerna från vinnarfamiljen ytterligare genom att skapa varje trovärdig modell med modulatoriska anslutningar, med tre begränsningar. För det första, både ansikten och objekt modulera minst en inter-areal anslutning. För det andra, vid Dubbelriktade länkar har de modulerande ingångarna en effekt på båda riktningarna. För det tredje, om face ger en direkt ingång till OFA antar vi att det alltid modulerar även OFA–ffa-anslutningen (Se tabell A2 i bilagan). Dessa modeller infördes i en andra familjevis slumpmässig BMS-analys., Slutligen, medlemmar av vinnaren underfamiljen in i en tredje BMS för att hitta en enda modell med den högsta överskridandesannolikheten.
resultat
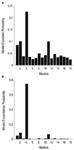
Bayesiansk modellval användes för att bestämma vilken modellfamilj som förklarar de uppmätta uppgifterna bäst. Som våra resultat visar den tredje familjen out-utförde de andra två, med en överskridande sannolikhet på 0.995 jämfört med den första familjens 0.00 och den andra familjens 0.004 (figurerna 3A,B)., Vinnaren modell familj (familj 3; Figur 2, botten) innehåller 12 modeller, som har kopplingar mellan LO och FFA, OFA och FFA, och LO och OFA samt, men skiljer sig i riktning av anslutningarna samt i stället för inmatning till nätverket.

Figur 3. Resultat av BMS RFX på nivån av familjer. A) de förväntade sannolikheterna för familjebaserad jämförelse visas, med de gemensamma sannolikheterna för överskridande (B).,
som ett andra steg konstruerades alla möjliga modulatoriska modeller för de 12 modellerna av familj 3 (Se tabell A2 i bilagan; material och metoder för detaljer). Detta ledde till 122 modeller, som ingick i BMS family-wise random analysis, med 12 underfamiljer. Som det är synligt från figurerna 4A, B, model sub-family 4 out-utförde de andra underfamiljerna med en överskridande sannolikhet på 0,79. Som ett tredje steg, var modellerna inom vinnaren underfamiljen 4 in i en slumpmässig effekt BMS. I Figur 5 presenteras de 18 testade variationerna av modell 20., Modellen med den högsta sannolikheten för överskridande (p = 0,75) var modell 4 (Se figur 6). Detta innebär att vinnaren modellen innehåller Dubbelriktade anslutningar mellan alla områden och ansikte och objekt ingångar, överraskande, båda in i LO. Dessutom har ansikten en modulerande effekt på sambandet mellan LO och FFA, medan objekt modulerar sambandet mellan LO och OFA.

Figur 4. Resultat av BMS RFX på nivån av underfamiljer., A) de förväntade sannolikheterna för familjebaserad jämförelse visas, med de gemensamma sannolikheterna för överskridande (B).

Figur 5. Modeller av underfamilj 4. Alla modeller har samma DCM.En struktur (identisk med modell 20 i Familj 3), DCM.B-strukturen skiljer sig från modell till modell. Streckade pilar symboliserade ansikte modulerande effekt, medan prickade fyrkantiga pilar visar objektmodulering.
Figur 6., Resultat av BMS inom underfamiljen 4. A) de förväntade sannolikheterna för familjebaserad jämförelse visas, med de gemensamma sannolikheterna för överskridande (B).
för att analysera parameteruppskattningar av vinnarmodellen över gruppen av ämnen användes en slumpmässig effekt approximation. Alla ämnesspecifika maximala a posteriori (karta) uppskattningar infördes i en t-test för enstaka medel och testades mot 0 (Stephan et al., 2010; Desseilles et al., 2011). Resultaten indexerade med en asterisk i Figur 7 skilde sig avsevärt från 0 (p < 0.,05).
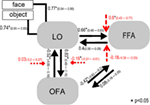
Figur 7. Strukturen av vinnaren modellen. Enkla linjer betecknar objektet och ansiktsstimuli till systemet (DCM.C). Svarta pilar visar interregionala anslutningar (DCM.A) medan de röda pilarna står för de modulerande anslutningarna (DCM.B): ansiktsmodulering är betecknad med streckade pilar, medan objektmodulering är betecknad med fyrkantiga Huvud streckade pilar. Medelvärden på gruppnivå för KARTBERÄKNINGAR och 95% konfidensintervall illustreras., Medelvärdena testades mot 0 och signifikanta resultat betecknas med * om p < 0.05.,
diskussion
det stora resultatet av den nuvarande effektiva anslutningsstudien tyder på att (A) LO är kopplad direkt till OFA–ffa-ansiktsbehandlingssystemet via Dubbelriktade anslutningar till båda områdena; (b) icke-ansikts-och ansiktsingångar är blandade i nivå med occipito-temporala områden och går in i systemet via LO; (C) ansiktsingång har en modulerande effekt på LO-och FFA-anslutningen, medan objektingången modulerar LO-och och en anslutning betydligt.
LO: s roll i objektuppfattning är välkänd från tidigare studier (Malach et al.,, 1995; Grillspector m.fl., 1998a, b, 2000; Lerner m.fl., 2001, 2008). Men trots tidigare studier fann vanligtvis en ökad aktivitet för komplexa föremål, liksom för ansikten i LO, är området vanligtvis förknippat med föremål och relativt mindre betydelse tillskrivs dess roll vid Ansiktsbehandling. I denna studie positionerade effektiv anslutningsanalys LO i kärnnätet för ansiktsuppfattning. Den direkta kopplingen mellan OFA och FFA har tidigare bevisats både funktionellt och anatomiskt (gschwind et al., 2012)., Eftersom det inte finns några aktuella uppgifter om LO: s roll i detta system har vi dock kopplat det till de två andra regionerna på flera rimliga sätt.
de första slumpmässiga BMS visade att familjen lo i winner-modellen är sammankopplad med både FFA och OFA och anslutningarna är dubbelriktade. Därför betonas att LO kan ha en direkt strukturell anslutning till FFA., Med modellering alla möjliga modulatoriska effekter fann vi att en underfamilj av modeller vann, där både objekt och ansikte ingångar in i systemet via LO, antar att LO spelar en allmän och viktig input region Roll. Sedan tidigare funktionella anslutningsstudier började alla analysen av ansiktsbehandlingsnätet på IOG-nivå, motsvarande OFA (Fairhall och Ishai, 2007; Ishai, 2008; Cohen Kadosh et al., 2011; Dima m.fl., 2011; Foley et al., 2012) Det är inte förvånande att de förbisett LO: s viktiga roll., Ansikten är dock faktiskt en distinkt kategori av visuella objekt, vilket tyder på att neuroner som är känsliga för föremål och former ska aktiveras, åtminstone i viss utsträckning, av ansikten också. Faktum är att encellsstudier av icke-mänskliga primater tyder på att den underlägsna temporala cortexen, den föreslagna homolog av humant LO i makakhjärnan (Denys et al., 2004; Sawamura et al., 2006) har neuroner som reagerar på ansikten också (Perrett et al., 1982, 1985; Desimone m.fl. 1984; Hasselmo m.fl., 1989; Unga och Yamane, 1992; Sugase et al., 1999)., Den intima anslutningen av LO till OFA och FFA, som föreslagits av den nuvarande studien, skulle kunna understryka det faktum att ansikten och föremål inte behandlas helt separat i den ventrala visuella vägen, en slutsats som stöds av de senaste funktionella bilddata också (Rossion et al., 2012). Den modulerande effekten av ansiktsinmatning på Lo-FFA-anslutningen tyder på att LO måste spela en roll vid Ansiktsbehandling, troligen kopplad till den tidigare strukturella behandlingen av ansikten, en uppgift som tidigare lagts ner mestadels till OFA (Rotshtein et al., 2005; Fox et al., 2009)., Slutligen visar gruppnivåparametrarna för vinnarmodellen att länken mellan LO, OFA och FFA huvudsakligen är hierarkiska och återkopplingarna från FFA mot OFA och LO spelar en svagare roll i systemet, åtminstone under den applicerade fixeringsuppgiften.
Sammanfattningsvis, genom att modellera den effektiva förbindelsen mellan ansiktsrelevanta områden och LO föreslår vi att LO spelar en viktig roll vid behandling av ansikten.,
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
bekräftelse
detta arbete stöddes av Deutsche Forschungsgemeinschaft (KO 3918/1-1) och Regensburgs universitet. Vi vill tacka Ingo Keck och Michael Schmitgen för diskussion.
Campbell, R. (2011). Speechreading och Bruce-Young-modellen för ansiktsigenkänning: tidiga resultat och den senaste utvecklingen. Br. J. Psychol. 102, 704–710.,
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Fairhall, S. L. och Ishai, A. (2007). Effektiv anslutning inom det distribuerade kortikala nätverket för ansiktsuppfattning. Cereb. Cortex 17, 2400-2406.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Hoffman, E. A., och Haxby, J. V. (2000). Distinkta representationer av ögon blick och identitet i det distribuerade mänskliga neurala systemet för ansiktsuppfattning. Nat. Neurosci. 3, 80–84.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Kourtzi, Z., och Kanwisher, N. (2001). Representation av uppfattad objektform av det mänskliga laterala occipitala komplexet. Vetenskap 293, 1506-1509.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
MacKay, D. J. C. (2003). Informationsteori, inferens och inlärningsalgoritmer. Cambridge: Cambridge University Press.
Nasanen, R. (1999). Rumslig frekvensbandbredd som används vid erkännande av ansiktsbilder. Vision Res.39, 3824-3833.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Rossion, B., Schiltz, C. Och Crommelinck, M., (2003b). Den funktionellt definierade höger occipital och fusiform ”ansiktsområden” diskriminerar roman från visuellt bekanta ansikten. Neuroimage 19, 877-883.
CrossRef fulltext
Sawamura, H., Orban, G. A. och Vogels, R. (2006). Selektivitet av neuronal anpassning matchar inte responselektivitet: en encellsstudie av fMRI-anpassningsparadigmet. Neuron 49, 307-318.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
















Lämna ett svar