Innledning
Chloroplasts representerer en strukturell funksjon av plante-celler og støtte plante overlevelse via deres primære stoffskifte og høyt nivå funksjoner (Kirk og Tilney-Bassett, 1978; Mulle, 1988; López-Juez og Pyke, 2005). I løpet av anlegget vegetativ vekst, blad celler inneholder en svært homogen befolkning på chloroplasts med hensyn til størrelse og form., Antall chloroplasts per celle er oppnådd ved binær fisjon av pre-eksisterende organeller og oppdeling i to datterceller under celledeling (Birky, 1983; Possingham og Lawrence, 1983). Dermed, regulering av chloroplast nummer i en gitt celle-type er avgjørende for cellenes funksjon og genetisk arv av chloroplasts.
for Å undersøke innholdet av chloroplast antall bestemmelse i plante-celler, blad mesophyll celler av representative arter har spilt en viktig rolle ., Disse cellene er fysiologisk viktig for fotosyntesen, og viser en høy grad av strukturell og funksjonell homogenitet. Tidlig systematisk observasjon analyser av isolert vev og celler (Boasson og Laetsch, 1969; Possingham og Saurer, 1969; Boffey et al., 1979; Lamppa et al., I 1980, Tomas og Rose, 1983; Pyke og Igle, 1991) gitt mye nyttig informasjon på chloroplast antall besluttsomhet, inkludert den oppfatningen at chloroplasts (plastids) er ikke syntetisert de novo men replikere ved divisjon og observasjon som leaf mesophyll chloroplast nummer er følsomme for ulike miljø-og plante-endogene faktorer. Med hensyn til sistnevnte, i spinat, lys har en positiv innvirkning på chloroplast divisjon i løpet blad plate kultur sammenlignet med dårlige lysforhold eller mørke forhold (Possingham og Lawrence, 1983)., I de første bladene av hvete, celle volum er positivt korrelert med chloroplast spredning (Ellis og Igle, 1985; Pyke og Igle, 1987). I Arabidopsis, den genetiske bakgrunnen påvirker chloroplast spredning; gjennomsnittlig chloroplast antall per celle i første bladene er 121 i Landsberg erecta (Ler) ecotype og 83 i Wassilewskija (Ws) ecotype (Pyke og Igle, 1994; Pyke et al., 1994)., Blad mesophyll celler har også bidratt til å forstå den genetiske kontroll av chloroplast divisjon; for eksempel screening mutanter svekket i chloroplast spredning og karakterisering av genet funksjoner som er involvert i chloroplast divisjon har avslørt at over 20 genene som koder chloroplast divisjon maskiner komponenter eller chloroplast regulerende faktorer (Gao og Gao, 2011; Miyagishima et al., 2011; Basak og Møller, 2013; Osteryoung og Pyke, 2014; Li et al., 2017).
i motsetning studier på replikering av chloroplasts i ikke-mesophyll celler (f.eks.,, fortau cellene i bladet epidermis; Itoh et al., 2018) er knappe. Nylig, regulering av chloroplast divisjon har blitt rapportert å variere mellom blad vev (Fujiwara et al., 2018; Itoh et al., 2018), selv om de detaljerte mekanismen er fortsatt ukjent. I tillegg, mens analyser av suspensjon-kultivert AV-2-celler og blad mesophyll protoplasts i tobakk og skyte apical meristem og blad opprinnelig celler i Arabidopsis (Nebenführ et al. I 2000; Sheahan et al.,, 2004; Seguí-Simarro og Staehelin, 2009) har gitt større innsikt, hvordan chloroplast (plastid) partisjonering er regulert i planter, er fortsatt uklart. Dermed, til tross for stor innsats, grunnleggende spørsmål i chloroplast forskning gjenstår, slik som (i) hvordan er chloroplast antall per celle koordinert i anlegget vev og (ii) hvordan er chloroplast partisjonering regulert ved celledeling.,
Historie av Forskning på Vakt Celle Chloroplast Antall
Stomatal GCs i skyte epidermis inneholder vanligvis chloroplasts og kontroll gassutveksling mellom blad mesophyll og atmosfæren (Sachs, 1875; Taiz et al., 2015, se Figur 1A). Den første undersøkelsen av GC chloroplast nummer i bladene ble utført i løpet av et århundre siden, i naturlig dyrket Drosera planter (Macfarlane, 1898)., Denne studien viste at i likhet med andre anlegg og celle strukturelle funksjoner, GC chloroplast antall per celle i et antatte hybrid stammer fra en krysning mellom Drosera filiformis og Drosera intermedia ble mellomting mellom de to artene, noe som tyder på at GC chloroplast nummeret kan brukes til å bestemme den genetiske sammensetningen av en plante. Viktige observasjoner ble senere rapportert om forskjeller i GC chloroplast antall blant plantearter (Sakisaka, 1929) og den relativt stabil chloroplast nummer i GCs i bladet epidermis av morbær (Morus spp.,; Hamada og Baba, 1930) og i modne blader av flere Brassica-arter (Iura, 1934). Videre analyse av autopolyploid sukker bete (Beta vulgaris) planter åpenbart at GC chloroplast nummer i blader er positivt korrelert med kjernefysiske ploidy nivå av planter (Mochizuki og Sueoka, 1955). Mer i dybden og omfattende analyser ble deretter gjennomført ved hjelp av ulike plante-prøver for å undersøke forholdet mellom chloroplast antall og stomatal størrelse med ploidy nivå (f.eks., Frandsen, 1968)., I disse analysene, chloroplast telle på stomatal (GC-par) nivå ofte ble vedtatt, som utelukket effekten av partisk chloroplast fordelingen mellom de sammenkoblede GCs (f.eks., Mochizuki og Sueoka, 1955; Frandsen, 1968), og viser at den gjennomsnittlige GC chloroplast nummer i blader eller cotyledons i ca 80 arter, varianter, eller hybrider varierte fra 2,8 til 40.0 i diploids (2×) og 5.0 til 73.5 i tetraploids (4×). I tillegg, hel-genom duplisering hendelser i planter (dvs., 1× 2×, 2× 4×, etc.) forårsaket en ca 1.,7-fold økning i GC chloroplast antall med high fidelity (anmeldt i Butterfass, 1973). Disse resultatene oppmuntret undersøkelser ploidy nivå i ulike vev og planter som er oppnådd via vev kultur, veikryss, eller naturlig dyrking, i kombinasjon med kjemiske (f.eks., colchicine) eller strålebehandling (for eksempel, Jacobs og Yoder, 1989; Singsit og Veilleux, 1991; Qin og Rotino, 1995)., Mens GC chloroplast nummer har blitt studert i stomatal biologi (Lawson, 2009) og cytologi å forstå chloroplast multiplikasjon (Butterfass, 1979; se nedenfor), er det i stor grad har fungert som en pålitelig og praktisk markør for påvisning av hybrider, arter og varianter, og for estimering av ploidy nivåer av mål-anlegg vev.
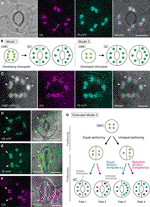
Figur 1 Forenklet modeller og mikroskopi bevis for kontroll av chloroplast nummer i stomatal vakt celler (GCs)., (A) En vanlig stomi (GC-par) i abaxial overhuden av Arabidopsis blad blad å uttrykke en stomi-målrettet fusjon av transitt-peptid (TP) med cyan fluorescerende protein (CFP; TP-CFP). (B) To modeller av chloroplast antall bestemmelse i GCs, som involverer enten chloroplast partisjonering (modell 1) eller begge chloroplast spredning og partisjonering (modell 2) ved GC-utvikling fra guard mor celler (GMCs). (C) EN GC-par i adaxial epidermis av Arabidopsis blad petiole å uttrykke TP-CFP og FtsZ1 smeltet til grønt fluorescerende protein (GFP; FtsZ1-GFP)., (D–F) GC-par i abaxial epidermis av Arabidopsis blad blad med (D, E) eller uten (F) uttrykk for TP smeltet sammen til den gule fluorescerende protein (YFP; TP-YFP). (F) Klorofyll autofluorescence (Chl) ble brukt som en chloroplast markør., (G) Utvidet modell 2, som representerer involvering av lik og ulik chloroplast partisjonering følgende GMC divisjon og påfølgende delingen av GC chloroplasts med lik (blå linje) eller selektivt (rød linje) divisjon kompetanse, noe som ville resultere i fire typer chloroplast antall bestemmelse (Skjebner 1-4) i løpet av sen stomatal utvikling av Arabidopsis blader. (A, C–F) Epifluorescence mikroskopi ble utført med et Olympus IX71 invertert mikroskop ved hjelp av plante-materialer som beskrevet tidligere (Fujiwara et al., 2017, Fujiwara et al., 2018)., Fluorescens-signaler av klorofyll, CFP, GFP, og YFP er pseudo-farget i magenta, cyan, grønn (i flettede bildet), og grønn, henholdsvis. Indikasjoner i paneler er som følger: arrowhead, i FtsZ1 ringen, pil, forstørret GC chloroplast; stiplede linjen, celle form. Målestokk = 10 µm.
Utility av Blad Vakt Celler For Analyse Av Chloroplast Antall Kontroll
Blad mesophyll celler har lenge blitt brukt som en primær modell for analyse av chloroplast antall., Mens de har fordeler for studier av virkninger av miljøforhold på chloroplast divisjon (f.eks., lys-cytokinin signaliserer; Boasson og Laetsch, 1969; Possingham og Lawrence, 1983; Okazaki et al., 2009; Chiang et al., 2012), de er begrenset i noen henseender. For det første, blad mesophyll celler som varierer i størrelse og form og er fordelt dypt inne i bladet, noe som gjør det vanskelig å manipulere intakt vev., For det andre, mottakelighet av blad mesophyll chloroplast spredning til miljømessige stress og plantevekst forhold kan hindre pålitelige sammenligninger mellom studier. Bladet mesophyll chloroplast antall per celle i Arabidopsis Columbia (Col) ecotype har blitt rapportert blant annet som 76 (Frende og Pyke, 1998), 80-100 (Stokes et al., 2000), 70 (Tirlapur og König, 2001), 41 (Han et al., 2007), 40-60 (Okazaki et al., 2009), og 30-40 (Kawade et al., 2013)., Videre, det er nesten umulig å vurdere bidraget fra chloroplast partisjonering til siste chloroplast antall per mesophyll celle i bladet utvikling, selv om dette er tenkt å være bestemt av balansen mellom frekvensen av celledeling og pris av chloroplast divisjon. For å avdekke mekanismen av chloroplast nummer kontroll i vegetativ blad celler, en modell system som overvinner problemene ovenfor er nødvendig.
Stomatal GCs (se Figur 1A) viser egenskapene til en modell system for å forstå mekanismen av chloroplast antall kontroll., GCs er svært jevn i størrelse og form i et vev, og deres spredt, men tett distribusjon i det ytterste laget av skudd forenkler deres deteksjon av lys-og fluorescens mikroskopi. GCs er også avledet fra protodermal celler i skyte apical meristem eller fra embryonale epidermal celler, og deres utviklingsmessige sekvens gjennom meristemoids (en stomatal forløper med meristematic aktivitet) og vakt mor celler (GMCs; en forløper av GC-par) er etablert i detalj (Zhao og Sekk, 1999; Nadeau og Sekk, 2002; Kalve et al., 2014)., Sent stomatal utvikling innebærer en enkelt runde av symmetriske GMC divisjon, som gjør det mulig vurdering av chloroplast distribusjon og partisjonering før og etter cytokinesis. Fra perspektivet av praktiske eksperimenter, blad GCs er egnet for mikroskopi. Det ble tidligere vist at chloroplast antall per celle i blad GCs av Sinapis alba var mindre påvirket av ulike lysforhold enn i blad mesophyll celler (Vill og Ulv, 1980)., I tillegg forskjellen i GC chloroplast nummer i blad petioles er relativt små mellom de tre Arabidopsis ecotypes Col, Ler, og Ws (Fujiwara et al., 2018). Videre, endoreduplication, som påvirker utviklingen av blad mesophyll, fortau, og trichome celler, har ikke vært påvist i Arabidopsis blad GCs (Melaragno et al., 1993), noe som vil sikre tolkninger av chloroplast antall data på 2C nivå av celler., Sammen disse rapportene tyder på at leaf GCs er potensielt en utmerket modell for systematisk analyse av chloroplast antall dynamikk i en bestemt celle avstamning.
Arabidopsis Blad Vakt Celler som en Modell for å Studere Kontroll av Chloroplast Antall
I historien av GC chloroplast forskning, chloroplast telle på stomatal (GC-par) nivå har vært en like viktig rolle i å bestemme chloroplast antall som teller på den enkelte GC-nivå. Begge metoder kan produsere det samme mener chloroplast antall (Butterfass, 1973)., Når variasjonen i chloroplast distribusjon i sammenkoblede GCs og dens underliggende mekanismen er gjenstand for fokus, detaljert informasjon om chloroplasts på den enkelte celle-nivå, dvs., størrelse, form, og intracellulære lokalisering, er viktig. Chloroplast (plastid) spredning i GMC–GC differensiering var tidligere undersøkt i flere plantearter (Butterfass, 1973, Butterfass 1979)., I disse studiene ble det foreslått to modeller for fastsettelse av terminal chloroplast nummer i GCs i forskjellige plantearter (Figur 1B): en (modell 1; sukkerroer) innebærer bare chloroplast oppdeling i GMC-divisjon, og det andre innebærer ikke bare chloroplast partisjonering, men også chloroplast spredning i løpet av GC-utvikling.
I den æra av molekylær genetikk, genomics, cell imaging, og andre tverrfaglige analyser, er det mange muligheter for videre karakterisering av chloroplast partisjonering mekanisme., Arabidopsis blad GCs kan være en av de beste modellen systemer for dette formålet. Flere studier har undersøkt GC chloroplast nummer i blader eller cotyledons av Arabidopsis (Hoffmann, 1968; Pyke og Igle, 1994; Pyke et al., 1994; Robertson et al. I 1995; Keech et al., 2007; Chen et al., 2009; Yu et al., 2009; Higaki et al., 2012; Fujiwara et al., 2018). Disse GCs viser et beskjedent antall chloroplasts, alt fra 3,5 til 5,5 i gjennomsnitt. Til dags dato, ingen studier har undersøkt endringer i chloroplast (plastid) antall under stomatal utvikling., Imidlertid, mikroskopiske bevis fra stomatal utvikling analyser (f.eks., Zhao og Sekk, 1999; Hachez et al. 2011) og vår foreløpige observasjoner tyder på at GMCs kan inneholde mindre tall for å utvikle chloroplasts enn GCs og at chloroplast spredning kan oppstå i løpet av GC differensiering. For å teste dette, dannelsen av chloroplast divisjon maskiner i GCs ble overvåket med sonden FtsZ1 smeltet til grønt fluorescerende protein (FtsZ1-GFP) (Fujiwara et al., 2008)., En transgene linjen, samtidig som uttrykker en transitt peptid (TP)-smeltet CFP og FtsZ1-GFP å visualisere stroma og FtsZ1 ringen, henholdsvis, ble undersøkt av epifluorescence mikroskopi (Fujiwara et al., 2017). Utvide blad petioles (femte blader av 4-ukers gamle frøplanter) ble ansatt. Som et resultat, GCs med symmetrisk sammentrekkende chloroplasts ble funnet (Figur 1C). Disse chloroplasts dannet FtsZ1 ring, en chloroplast divisjon ring på stromal overflaten av den indre konvolutt membran i blad mesophyll og fortau celler (Vitha et al. I 2001; Fujiwara et al.,, 2008), i ekvatorial-innsnevring nettstedet. I samsvar med stomatal patterning i Arabidopsis blad utvikling (Donnelly et al. I 1999; Andriankaja et al., 2012), dele chloroplasts ble oppdaget på slutten, så vel som tidlig stadium av blad-utvidelsen. Dermed, modell 2 er mest sannsynlig den som passer best for Arabidopsis blad GCs.
Forholdet Mellom Chloroplast Spredning Og Ekspansjon i Arabidopsis Blad Vakt Celler
Videre, en uventet fenotypen av GC chloroplast morphogenesis ble observert i modne GCs (Figur 1D, E)., Når epidermal skreller av fullt utvidet blader (tredje–fjerde blad blader av 4-ukers gamle frøplanter) fra en TP-smeltet gule fluorescerende protein (YFP) linje var mikroskopisk preget (FL6-5 line; Fujiwara et al., 2018), noen stomata viste ulik chloroplast distribusjon mønstre i GC-par, mens de fleste blad stomata viste lik eller lignende chloroplast distribusjon mønstre (Robertson et al. I 1995; Fujiwara et al., 2018)., Innen GC-par i en stomi, størrelsen på chloroplasts i GC-som inneholder mindre tall av chloroplasts var større enn i de andre GCs i paret som inneholder et større antall chloroplasts (Figur 1D, E). På denne måten, GCs sannsynligvis opprettholde den totale chloroplast volum per celle på et konstant nivå i løpet av cellevekst. Forstørret chloroplasts representert terminalen fenotypen og ikke lenger kunne dele inn i utvidet blader. Disse resultatene ble bekreftet i flere uavhengige eksperimenter, uavhengig av uttrykk for en TP-smeltet fluorescerende protein for stroma merking (Figur 1F).,
Dette GC chloroplast fenotypen er tolket som en erstatning mekanisme for chloroplast ekspansjon, som ble godt dokumentert i blad mesophyll celler med defekt i kontroll av chloroplast divisjon (Pyke og Igle, 1994; Pyke et al., 1994). Til dags dato, kun en studie (Ellis og Igle, 1985) har rapportert en negativ korrelasjon mellom chloroplast antall og chloroplast størrelse i blad mesophyll celler av hvete, mens mange studier har rapportert en positiv sammenheng mellom celle volum og chloroplast nummer i normal blad mesophyll celler (Igle og Pyke, 1988; Pyke, 1997)., Mens ubalanser i GC chloroplast antall skje ved lave frekvenser (Fujiwara et al., 2018), chloroplast mangfold i GC-par indikerer at ulike chloroplast partisjonering kan utløse differensial chloroplast vekst mellom vill-type blad celler i Arabidopsis, til tross for symmetriske celledeling.
chloroplast kompensasjon effekt i GCs kan være mindre strenge enn i blad mesophyll celler., GCs kan være i stand til å tåle knapphet eller fullstendig tap av den totale chloroplast volum per celle i alvorlig nedsatt chloroplast divisjon mutanter, for eksempel i Arabidopsis arc6 og atminE1 og tomat suffulta, mens mange mutant GCs viste redusert chloroplast antall og utvidet chloroplast størrelse på samme måte til bladet mesophyll celler (Robertson et al. I 1995; Frem og Pyke, 2006; Chen et al., 2009; Fujiwara et al., 2018)., I en sen chloroplast divisjon mutant, arc5, reduksjon i GC chloroplast antall ikke var assosiert med en betydelig økning i chloroplast størrelse, i motsetning til i blad mesophyll celler (Pyke og Igle, 1994). En lavere grad av chloroplast ekspansjon i GCs enn i mesophyll celler (Pyke og Igle, 1994; Barton et al., 2016), og variasjonen i chloroplast ekspansjon blant GCs, kan ligge til grunn for et så bredt tillatt spekter av totalt chloroplast volum per GC., Videre er tidspunktet for chloroplast divisjon i løpet GMC–GC differensiering kan påvirke terminal GC chloroplast fenotypen. Selv om mer detaljert karakterisering er nødvendig for å løse dette problemet, og det virker sannsynlig at Arabidopsis blad GCs representerer et system for å undersøke uutnyttede aspekter av chloroplast nummer kontroll i plante-celler.,
En Fungerende Modell For Chloroplast Antall Bestemmelse i Arabidopsis Blad Vakt Celler
På grunnlag av ovennevnte, foreslår vi en fungerende modell (en utvidet modell 2) for analyse av chloroplast nummer i GCs (Figur 1). Den endelige chloroplast antall per GC er bestemt av chloroplast oppdeling i GMC divisjon og chloroplast spredning i GCs. I løpet av GMC divisjon, chloroplasts kan gjennomgå enten lik eller ulik oppdeling. Under chloroplast spredning, GC chloroplasts vil spre seg med enten lik (blå linje) eller selektivt (rød linje) divisjon kompetanse., For eksempel, hvis like partisjonert chloroplasts har tilsvarende divisjon kompetanse, lik chloroplast tall vil skje i GC-par (Skjebnen 1). Hvis ulikt partisjonert chloroplasts har tilsvarende divisjon kompetanse, chloroplasts vil øke i samme takt i GC-par (Skjebne 3). Hvis selektiv chloroplast divisjon oppstår i GCs, balansen av chloroplast nummer i GC-par vil endre seg etter GMC divisjon (Skjebner 2 og 4)., Det er for tiden vanskelig å finne støtte for «selektiv chloroplast divisjon,» men hvis Skjebner 1 og 4 er faktisk i flertall i GCs, og da ville de ha en mekanisme som styrer total chloroplast volum per celle, som i blad mesophyll celler. Modellen reiser to problemstillinger: (i) Er GC chloroplasts riktig partisjonert til dattercellene, og hvordan de partisjon? Og (ii) er divisjon kompetanse av GC chloroplasts coordinately regulert?,
Angående problemet (jeg), om chloroplast arv oppstår ved tilfeldig fordeling av flere chloroplasts i cytoplasma eller ved positive chloroplast partisjonering mekanismen(e) som har vært en langvarig bekymring (Butterfass, 1969; Birky, 1983; Hennis og Birky, 1984; Nebenführ, 2007; Sheahan et al., 2016). Intriguingly, i Arabidopsis arc6, blad eller cotyledon GCs har null til tre chloroplasts, og i chloroplast-mangelfull GCs, ikke-fotosyntetiske plastids fortsatt eksisterer i vesicular til avlange former (Robertson et al. I 1995; Chen et al., 2009; Fujiwara et al., 2018)., Ingen GCs blottet for plastids per se som er blitt funnet i arc6, og ingen forklaring på dette har vært i møte, til tross for avbrudd av chloroplast divisjon apparater (Vitha et al., 2003). Følgelig vil det være viktig å undersøke replikering og morfologi av arc6 chloroplasts i stomatal avstamning studier. Arabidopsis mutant forskning kan også gi en annen ledetråd for dette problemet., Den observasjon at 18% av cotyledon GCs i krøllete blad (crl) mutant inneholder ingen plastidic strukturer i cytoplasma, mens 100% av blad mesophyll celler som inneholder én til fire forstørret chloroplasts (Asano et al., 2004; Chen et al., 2009), er av stor betydning. CRL er en chloroplast ytre-konvolutt protein med ukjent funksjon. Forståelse CRL protein funksjon kan gi innsikt i mekanismen(e) av chloroplast partisjonering., Analyse av chloroplast spredning og oppdeling i blad mesophyll celler i Arabidopsis arc mutanter og andre transgene linjer har fremmet forskning på spredning og oppdeling av ikke-mesophyll plastids. Likeledes, resultater oppnådd i GCs kan være overførbar til andre celle-systemer.
Siste Kommentar
GC-modellen åpner mange muligheter for utvikling av chloroplast biologi. For eksempel, mens cytoskeletal systemer er kjent for å regulere chloroplast morfologi, bevegelse og partisjonering (Sheahan et al.,, 2016; Wada, 2016; Eriksen og Schattat, 2018), rollen til hvert gen regulatoriske i chloroplast spredning og oppdeling i planter har fått lite oppmerksomhet. På den annen side, når det blir mulig å svekke GC chloroplast nummer eller morfologi via ulike eksperimentelle strategier, ny innsikt i de molekylære kontroll av chloroplast morphogenesis i stomatal avstamning celler kan bli gitt. I tillegg, i forbindelse med kvantitative analyser av chloroplast nummer i stomatal utvikling, matematisk modellering kan tilby en ny vei for disse undersøkelsene., Denne artikkelen presenterer aktuell kunnskap om hvordan GC chloroplast nummer er kontrollert og fremhever den potensielle nytten av Arabidopsis blad GCs for å forstå chloroplast spredning og partisjonering.
Data Tilgjengelighet Uttalelse
Alle datasett for denne studien er inkludert i artikkelen/ supplerende materiale.
Forfatter Bidrag
MF unnfanget studien og skrev manus. MF og SOM gjennomførte eksperimenter. SOM og RI utført analysene. Alle forfattere lest og godkjent det siste manuskriptet.,
Midler
Dette arbeidet ble støttet av Departementet for Utdanning, Kultur, Vitenskap og Teknologi i Japan under KAKENHI (grant nr. 19K05831 til MF og 18K06314 til RI).
interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller finansielle forhold som kan oppfattes som en potensiell interessekonflikt.
Erkjennelsene
forfatterne takke Dr. Nobuyuki Kanzawa (Sofia Universitet) for råd om arbeidet.
Birky, C. W., Jr., (1983)., Partisjonering av cytoplasmatiske dannes ved celledeling. Int. Rev. Cytol. 15, 49-86.
Google Scholar
smør churn, T. (1969). Den plastid distribusjon i mitose av den avsluttende celle mor celler av haploid swedenklee (Trifolium hybridum L.). Planta 84, 230-234. doi: 10.1007/BF00388108
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar
smør churn, T. (1973). Kontroll av plastid divisjon ved hjelp av kjernefysiske DNA-mengden. Protoplasmaen 76, 167-195. doi: 10.,1007/BF01280696
CrossRef Full Text | Google Scholar
Qin, X., Rotino, G. L. (1995). Chloroplast number in guard cells as ploidy indicator of in vitro-grown androgenic pepper plantlets. Plant Cell Tissue Organ Cult. 41, 145–149. doi: 10.1007/BF00051583
CrossRef Full Text | Google Scholar
Sachs, J. (1875). Textbook of Botany. Oxford: Clarendon Press.
Google Scholar
Thomas, M. R., Rose, R. J. (1983)., Plastid antall og plastid strukturelle endringer knyttet til tobakk mesophyll protoplast kultur og anlegg regenerering. Planta 158, 329-338. doi: 10.1007/BF00397335
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar
Tirlapur, U. K., König, K. (2001). Femtosecond nær-infrarøde lasere som en roman verktøy for ikke-invasive real-time high-resolution time-lapse avbildning av chloroplast divisjon i levende bunt skjede celler av Arabidopsis. Planta 214, 1-10. doi: 10.,1007/s004250100597
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar
Wild, A., Ulv, G. (1980). Effekten av forskjellige lys intensitet på hyppighet og størrelse av stomata, størrelsen på cellene, antall, størrelse og innhold av klorofyll chloroplasts i mesophyll og vakt celler under ontogeny av primære blader av Sinapis alba. Z. Pflanzenphysiol. Bd. 97, 325-342. doi: 10.1016/S0044-328X(80)80006-7
CrossRef Full Tekst | Google Scholar
















Legg igjen en kommentar