Indledning
Det neurale behandling af ansigter er et meget undersøgt emne kognitiv videnskab. Baseret på funktionelle billeddannelsesundersøgelser, enkeltcelleoptagelser og neuropsykologisk forskning er det blevet antydet, at Ansigtsbehandling udføres af et distribueret netværk, der involverer flere kortikale områder af pattedyrshjernen (Ha .by et al., 2000; Marotta et al., 2001; Rossion et al., 2003a; Avidan et al., 2005; Sorger et al., 2007)., Mens omfanget af dette ansigt-behandling netværk er i øjeblikket under intensiv debat (Ishai, 2008; Wiggett og Downing, 2008) de fleste forskere er enige om, at der er mange kortikale områder, der aktiveres af ansigt stimuli. De mest indflydelsesrige model af ansigt opfattelse, baseret på den oprindelige model af Bruce og Unge, 1986; Young og Bruce, 2011) foreslår en skelnen mellem repræsentation af størrelse og variant aspekter af ansigt opfattelse i en relativt uafhængig måde, som er opdelt i “kerne” og en “udvidet” del (Haxby et al., 2000)., De vigtigste regioner i “kernenetværket” er områder af occipital og den laterale fusiform gyri (FG). De områder, der er af disse to anatomiske regioner synes at være specialiserede til forskellige opgaver: mens occipital ansigtet område (OFA), som ligger på den ringere occipital gyrus (IOG) synes at være involveret i den strukturelle behandling af ansigter, den tenformede ansigtet område (FFA) processer ansigter på et højere niveau måde, der bidrager for eksempel til behandling af identitet (Sergent et al., 1992; George et al., 1999; Ishai et al., 1999; Hoffman og Ha Haby, 2000; Rossion et al., 2003a,b; Rotshtein et al.,, 2005). Derudover er de foranderlige aspekter af ansigter (såsom ansigtsudtryk, retning af øje–blik, udtryk, læbebevægelser (Perrett et al., 1985, 1990) eller læbe-læsning (Campbell, 2011) ser ud til at blive behandlet i den overlegne temporale sulcus (STS; Puce et al., 1998; Hoffman og Ha Haby, 2000; Hoffmaninston et al., 2004). De tre ovennævnte områder (FFA, OFA, STS) danner den såkaldte “kerne” af det perceptuelle system for Ansigtsbehandling (Ha .by et al., 2000; Ishai et al., 2005)., Mens grundlæggende oplysninger om ansigter behandles af dette kernesystem, tilføjer komplekse oplysninger om andres humør, interesseniveau, attraktivitet eller opmærksomhedsretning også information til ansigtsopfattelse og behandles af et yderligere, såkaldt “udvidet” system (Ha .by et al., 2000). Dette system indeholder hjerneområder med en lang række kognitive funktioner relateret til behandlingen af foranderlige ansigtsaspekter (Ha .by et al., 2000; Ishai et al., 2005) og inkluderer områder som amygdala, insula, den underordnede frontale gyrus såvel som den orbitofrontale Corte. (Ha .by et al.,, 2000, 2002; Fairhall og Ishai, 2007; Ishai, 2008).interaktioner mellem de ovennævnte områder modelleres i den foreliggende undersøgelse ved hjælp af metoder, der beregner den effektive forbindelse mellem kortikale områder. Dynamisk kausal modellering (DCM) er en meget anvendt metode til at udforske effektiv forbindelse mellem hjerneområder. Det er en generisk tilgang til modellering af den gensidige indflydelse fra forskellige hjerneområder på hinanden, baseret på fMRI-aktivitet (Friston et al., 2003; Stephan et al., 2010) og at estimere interforbindelsesmønsteret for kortikale områder., DCMs er generative modeller af neurale reaktioner, som giver “a posteriori” estimater af synaptiske forbindelser blandt neuronale populationer (Friston et al., 2003, 2007). Eksistensen af distribueret netværk for ansigt-behandling blev først bekræftet af funktionelle forbindelser analyse af Ishai (2008), der hævdede, at det centrale knudepunkt for ansigt-behandling er den laterale FG, som er forbundet til lavere for områder af IOG samt til STS, amygdala, og frontale områder. Det første forsøg på at afsløre ansigtsbehandlingsnetværket i tilfælde af realistiske dynamiske ansigtsudtryk blev udført af Foley et al. (2012)., De bekræftede rollen som iog -, STS-og FG-områder og fandt, at forbindelsesstyrken mellem medlemmer af kernenettet (OFA og STS) og det udvidede system (amygdala) øges til behandling af påvirkningsbelastede bevægelser. I øjeblikket uddybede flere undersøgelser vores forståelse på ansigtsbehandlingsnetværket og afslørede en direkte forbindelse mellem amygdala og FFA og denne forbindelses rolle i opfattelsen af frygtelige ansigter (Morris et al., 1996; Marco et al., 2006; Herrington et al., 2011)., Endelig blev effekten af højere kognitive funktioner på ansigtsopfattelsen også modelleret ved at teste forbindelserne mellem den orbitofrontale Corte.og kernenetværket (Li et al., 2010). Det blev fundet (Li et al ., 2010) at den orbitofrontale Corte.har en effekt på OFA, som yderligere modulerer informationsbehandlingen af FFA.mens tidligere effektive forbindelsesundersøgelser afslørede detaljerne i ansigtsbehandlingsnetværket relateret til forskellige aspekter af ansigtsopfattelse, ignorerede de den enkle kendsgerning, at ansigter også kan betragtes som visuelle objekter., Vi ved fra en stor mængde eksperimenter, at visuelle objekter behandles af et distribueret kortikalt netværk, herunder tidlige visuelle områder, occipito-temporal og ventral–temporal cortices, der stort set overlapper med ansigtsbehandlingsnetværket (Ha .by et al., 1999, 2000, 2002; Kourt Andi og kan Kanisher, 2001; Ishai et al., 2005; Gobbini og Ha andby, 2006, 2007; Ha .by, 2006; Ishai, 2008). En af de store områder af visuelle objekter, der skal forarbejdes, er den laterale occipital cortex (LOC), som kan opdeles i to dele: den anterior–ventrale (PF/LOa) og den caudale–dorsale del (LO; Grill-Spector et al.,, 1999; Halgren et al., 1999). LOC blev først beskrevet af Malach et al. (1995), der målte øget aktivitet for objekter, herunder også berømte ansigter, sammenlignet med krypterede objekter (Malach et al., 1995; Grill-Spector et al., 1998a). Siden da betragtes den laterale occipital (LO) primært som et objektselektivt område, som alligevel altid viser sig at have forhøjet aktivering for ansigter også (Malach et al., 1995; Puce et al., 1995; Lerner et al., 2001), især for inverterede (Aguirre et al., 1999; Ha ;by et al., 1999; Epstein et al.,, 2005; Yovel and kan .isher, 2005).
det er således temmelig overraskende, at selvom flere undersøgelser har behandlet den effektive forbindelse af ansigtsbehandlingsområder, betragtede ingen af dem LO ‘ s rolle i netværket. I en tidligere fMRI-undersøgelse fandt vi, at LO har en afgørende rolle i sensorisk konkurrence om ansigtsstimuli (Nagy et al., 2011)., Aktiviteten af LO blev reduceret ved præsentationen af samtidigt præsenterede samtidige stimuli, og denne responsreduktion, der afspejler sensorisk konkurrence blandt stimuli, var større, når den omgivende stimulus var et ansigt sammenlignet med et Fourier-fase randomiseret støjbillede. Dette resultat understøttede også ideen om, at LO kan spille en bestemt rolle i ansigtsopfattelsen. Derfor undersøgte vi eksplicit ved hjælp af metoder til effektiv forbindelse, hvordan LO er knyttet til FFA og OFA, medlemmer af det foreslåede kernenetværk for ansigtsopfattelse (Ha .by et al., 1999, 2000, 2002; Ishai et al., 2005).,
materialer og metoder
emner
femogtyve sunde deltagere deltog i eksperimentet (11 kvinder, median: 23 år, min.: 19 år, ma..: 35 år). Alle havde normal eller korrigeret til normal vision (selvrapporteret), ingen af dem havde neurologiske eller psykologiske sygdomme. Fag gav deres skriftlige informerede samtykke i overensstemmelse med de protokoller, der er godkendt af det etiske udvalg ved University of Regensburg.,
Stimuli
Emner, der var centralt præsenteret af grå-skala ansigter, ikke-følelse, objekter, og Fourier-randomiseret versioner af disse stimuli, skabt af en algoritme (Nasanen, 1999), som erstatter den fase spektrum med tilfældige værdier (fra 0° til 360°), forlader amplitude spektrum af billedet intakt, samtidig med at fjerne enhver form for information. Ansigter var fuldfront digitale billeder af 20 unge mænd og 20 unge kvinder. De var fit bag en rund form maske (3,5 diameter diameter) fjerne de ydre konturer af ansigterne (se et eksempelbillede i Figur 1)., Objekter var ikke-sense, gengivne objekter (n = 40) med samme gennemsnitlige størrelse som ansigtsmasken. Luminansen og kontrasten (dvs.standardafvigelsen for luminansfordelingen) af stimuli blev ligestillet ved at matche luminanshistogrammerne (gennemsnitlig luminans: 18 cd/m2) ved hjælp af Photoshop. Stimuli blev tilbage-projiceret via en LCD-videoprojektor (JVC, DLA-G20, Yokohama, Japan, 72 h., 800. 600 opløsning) på en gennemskinnelig cirkulær skærm (ca. 30 diameter diameter), placeret inde i scannerboringen 63 cm fra observatøren., Stimulus præsentation blev kontrolleret via E-prime soft .are (Psychological Soft .are Tools, Pittsburgh, PA, USA). Ansigter, objekter og Fourier-støjbilleder blev præsenteret i efterfølgende blokke på 20 s, interleaved med 20 s tomme perioder (ensartet grå baggrund med en luminans på 18 cd/m2). Stimuli blev præsenteret for 300 ms og blev efterfulgt af en isi på 200 ms (2 H.) i tilfældig rækkefølge. Hver blok blev gentaget fem gange. Deltagerne blev bedt om at fokusere kontinuerligt på et centralt præsenteret Fi markeringsmærke., Disse funktionelle locali .er-kørsler var en del af to andre eksperimenter med ansigtsopfattelse, der blev offentliggjort andetsteds (Nagy et al., 2009, 2011).

Figur 1. Prøve stimuli af eksperimentet. Alle billeder var gråskala, samme i størrelse, luminans og kontrast. Venstre panel viser et ansigt, kønsspecifikke funktioner (såsom hår, smykker osv.) var skjult bag en oval maske. Midterste panel viser en prøve ikke-sense geometrisk objekt, mens højre panel viser Fourier-fase randomiseret version af objekter, der anvendes som kontrol stimuli.,
dataindsamling og Analyse
Imaging blev udført ved hjælp af en 3-T HR. Hoved-scanner (Siemens Allegra, Erlangen, Tyskland). For funktionel serie, vi løbende har erhvervet billeder (29 skiver, 10° hælder i forhold til aksial -, T2 – * vægtet EPI sekvens, TR = 2000 ms; TE = 30 ms, flip angle = 90°; 64 × 64 matricer; i-fly opløsning: 3 mm × 3 mm; skive tykkelse: 3 mm). Sagittale T1-vægtede billeder i høj opløsning blev erhvervet ved hjælp af en magnetiseringsepi-sekvens (MP-RAGE; tr = 2250 ms; TE = 2.,6 ms; 1 mm isotrop vo .elstørrelse) for at opnå en 3D strukturel scanning (for detaljer, se Nagy et al., 2011).
Funktionelle billeder blev korrigeret for erhvervelsen forsinkelse, forlagte, normaliseret til MNI plads, resampled til 2 mm × 2 mm × 2 mm opløsning og rumligt glattes med en Gauss-kerne på 8 mm FWHM (SPM8, Welcome Department of Imaging Neuroscience i London, UK; for detaljer analyse af data, se Nagy et al., 2011).
VOI-valg
først blev interessevolumener (VOI) valgt, baseret på aktivitet og anatomiske begrænsninger (inklusive maskering for relevante hjerneområder)., Ansigtsselektive områder blev defineret som et område, der viste større aktivering for ansigter sammenlignet med Fourier-støjbilleder og objekter. FFA blev defineret inden for den laterale Fusiform Gyrus, mens OFA inden for IOG. LO blev defineret fra objektet > Fourier støj og ansigt billeder kontrast, inden for den midterste Occipital Gyrus. VOI-udvælgelsen var baseret på T-kontrasterne justeret med f-kontrast, (p < 0.005 ukorrigeret med en klyngestørrelse på mindst 15 vo .els)., VOIs var sfærisk med en radius på 4 mm omkring topaktiveringen (for individuelle koordinater, Se tabel A1 i Appendiks). Variansen forklaret af den første egenvariat af de dristige signaler var alle over 79%. Kun højre halvkugleområder blev brugt i den nuværende DCM-analyse, da flere undersøgelser peger på denne halvkugles dominerende rolle i ansigtsopfattelsen (Michel et al., 1989; Sergent et al., 1992).,
Effektiv Tilslutning Analyse
Effektiv forbindelse blev testet af DCM-10, der er gennemført i SPM8 toolbox (Wellcome Department of Imaging Neuroscience i London, england, STORBRITANNIEN), der kører under Matlab R2008a (The MathWorks, Natick, MA, USA). Modeller af DCM er defineret med endogene forbindelser, der repræsenterer koblingen mellem hjerne-regioner (matrix A), modulerende forbindelser (matrix B), og kørsel input (matrix C). Her i A-Matri theen definerede vi forbindelserne mellem de ansigtsselektive regioner (FFA og OFA) og LO., Billeder af ansigter og genstande tjente som køreindgang (Matri.C), og på dette analysetrin anvendte vi ingen modulatoriske effekter på forbindelserne.
model estimering til formål at maksimere de negative fri-energi estimater af modellerne (f) for et givet datasæt (Friston et al., 2003). Denne metode sikrer, at model fit bruger parametrene på en parsimonious måde (E .bank et al., 2011). De estimerede modeller blev sammenlignet, baseret på modellen evidences p(y|m), hvilket er sandsynligheden p for at opnå observerede data y givet af en bestemt model m (Friston et al., 2003; Stephan et al., 2009)., I den foreliggende undersøgelse anvender vi den negative fri-energi tilnærmelse (variational free-energy) til log beviser (MacKay, 2003; Friston et al., 2007). Bayesian Model Selection (BMS) blev udført på både tilfældige (RF.) og faste (ff.) effektdesign (Stephan et al., 2009). BMS RF.er mere modstandsdygtig over for outliers end FF., og det antager ikke, at den samme model ville forklare funktionen for hver deltager (Stephan et al., 2009). Med andre ord er RF.mindre følsom over for støj., I RF. – tilgangen er analysens output sandsynligheden for overskridelsen af modelrummet, hvilket er i hvilket omfang en model er mere tilbøjelig til at forklare de målte data end andre modeller. Den anden udgang af RF. – analysen er den forventede posterior sandsynlighed, hvilket afspejler sandsynligheden for, at en model genererede de observerede data, hvilket tillader forskellige distributioner til forskellige modeller. Begge disse parameterværdier reduceres ved at udvide modelrummet (dvs ., derfor opfører de sig på en relativ måde, og modeller med delte funktioner og usandsynlige modeller kan forvrænge analysens output. Ud over den direkte sammenligning af de 28 oprettede modeller delte vi derfor modelrummet i familier med lignende forbindelsesmønstre ved hjælp af Penny et al. (2010).
siden flere tidligere DCM-undersøgelser peger på den tætte tovejsforbindelse mellem FFA og OFA (Ishai, 2008; Gsch .ind et al.,, 2012) i vores Analyse var disse to områder altid knyttet til hinanden, og LO var forbundet med dem på enhver biologisk plausibel måde. De 28 relevante modeller blev opdelt i tre modelfamilier baseret på strukturelle forskelle (Penny et al., 2010; E .bank et al., 2011). Familie 1 indeholder modeller med lineære forbindelser mellem de tre områder, antager, at information strømmer fra LO til FFA via OFA. Familie 2 indeholder modeller med en trekantet konstruktion, hvor LO sender input direkte til FFA og OFA er også direkte knyttet til FFA., Familie 3 indeholder modeller, hvor de tre områder er indbyrdes forbundne, idet man antager en cirkulær informationsstrøm (figur 2). For at begrænse antallet af modeller i dette trin i analysen modulerede indgangene udelukkende aktiviteten i deres indgangsområder. De tre familier blev sammenlignet med et tilfældigt design BMS.
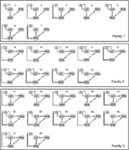
Figur 2. De 28 analyserede modeller. Sorte linjer markerer opdelingen mellem de tre familier, der har forskellige Matri structuresstrukturer. For detaljer, se afsnittet ” Materialer og metoder.,”
for det andet blev modellerne fra vinderfamilien uddybet yderligere ved at skabe enhver plausibel model med modulatoriske forbindelser og anvende tre begrænsninger. For det første modulerer både ansigter og objekter mindst en interregional forbindelse. For det andet, i tilfælde af tovejsforbindelser har de modulatoriske indgange en effekt på begge retninger. For det tredje, hvis ansigt giver et direkte input til OFA, antager vi, at det altid modulerer OFA–FFA-forbindelsen også (se tabel A2 i tillæg). Disse modeller blev indgået i en anden familievis tilfældig BMS-analyse., Endelig blev medlemmer af winnerinner-underfamilien indgået i en tredje BMS for at finde en enkelt model med den højeste overvægtssandsynlighed.
resultater
Bayesian modelvalg blev brugt til at afgøre, hvilken modelfamilie der forklarer de målte data bedst. Som vores resultater viser den tredje familie ud-udført de to andre, der har en overskridelse Sandsynlighed for 0,995 i forhold til den første familie 0,00 og den anden familie 0,004 (figur 3A,B)., Vinderen model familien (Familie 3; Figur 2, nederst) indeholder 12 modeller, der forbindelser mellem LO og FFA, OFA og FFA, og LO og OFA så godt, men forskellige i retning af de forbindelser, såvel som i stedet for input til netværket.

Figur 3. Resultater af BMS RF.på niveau med familier. (A) de forventede sandsynligheder for familiebaseret sammenligning er vist med de fælles overskridelsessandsynligheder (B).,
Som et andet trin, alle mulige modulerende modeller blev konstrueret til 12 modeller for Familie 3 (se Tabel A2 i Appendiks; Materialer og Metoder for detaljer). Dette førte til 122 modeller, der blev indgået i BMS-familiens tilfældige analyse ved hjælp af 12 underfamilier. Som det fremgår af figur 4A, B, udførte modelunderfamilien 4 de øvrige underfamilier med en overvægtssandsynlighed på 0,79. Som et tredje trin blev modellerne i winnerinner-underfamilien 4 indgået i en tilfældig effekt BMS. Figur 5 præsenterer de 18 testede variationer af Model 20., Modellen med den højeste sandsynlighed for overskridelse (p = 0,75) var model 4 (Se figur 6). Dette betyder, at vindermodellen indeholder tovejsforbindelser mellem alle områder og ansigts-og objektindgange, overraskende, begge går ind i LO. Derudover har ansigter en modulerende effekt på forbindelsen mellem LO og FFA, mens objekter modulerer forbindelsen mellem LO og OFA.

Figur 4. Resultater af BMS RF.på niveau med underfamilier., (A) de forventede sandsynligheder for familiebaseret sammenligning er vist med de fælles overskridelsessandsynligheder (B).

Figur 5. Modeller af underfamilie 4. Alle modeller har samme DCM.En struktur (identisk med Model 20 i familie 3), DCM.B struktur adskiller sig fra model til model. Stiplede pile symboliseret ansigt modulerende effekt, mens stiplede firkantede pile viser objekt modulation.
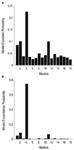
Figur 6., Resultater af BMS inden for underfamilien 4. (A) de forventede sandsynligheder for familiebaseret sammenligning er vist med de fælles overskridelsessandsynligheder (B).
til analyse af parameterestimater af vindermodellen på tværs af gruppen af forsøgspersoner blev der anvendt en tilfældig effekt tilnærmelse. Alle de fagspecifikke maksimale a posteriori (kort) estimater blev indgået i en t-test for enkeltmidler og testet mod 0 (Stephan et al., 2010; Desseilles et al., 2011). Resultaterne indekseret med en stjerne på figur 7 adskiller sig væsentligt fra 0 (p < 0.,05).
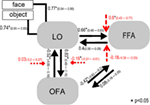
Figur 7. Strukturen af vinderen model. Enkle linjer betyder objektet og ansigt input stimuli til systemet (DCM.C). Sorte pile viser interregionale forbindelser (DCM.A) mens de røde pile står for de modulatoriske forbindelser (DCM.B): face modulation er betød med stiplede pile, mens objekt modulation er betød med firkantede-head stiplede pile. Gruppe-niveau gennemsnit af kort skøn og 95% konfidensintervaller er illustreret., Gennemsnittet blev testet mod 0, og signifikante resultater betegnes med * if p < 0.05.,
Diskussion
Det vigtigste resultat af de nuværende effektive tilslutningsmuligheder undersøgelsen tyder på, at (a) LO er knyttet direkte til OFA–FFA ansigt-system via tovejs forbindelser til begge områder; (b) ikke-face og face-indgange er blandet på niveau med occipito-temporale områder og komme ind i systemet via LO; (c) pålydende input har en modulerende effekt på LO og FFA-forbindelse, mens objekt input modulerer LO og OFA forbindelse væsentligt.
LO ‘ s rolle i objektopfattelse er velkendt fra tidligere undersøgelser (Malach et al.,, 1995; Grill-Spector et al., 1998a, B, 2000; Lerner et al., 2001, 2008). På trods af, at tidligere undersøgelser normalt fandt en øget aktivitet for komplekse objekter såvel som for ansigter i LO, er området normalt forbundet med objekter, og relativt mindre betydning tilskrives dets rolle i Ansigtsbehandling. I den foreliggende undersøgelse placerede effektiv forbindelsesanalyse LO i kernenetværket af ansigtsopfattelse. Den direkte forbindelse mellem OFA og FFA har tidligere vist sig både funktionelt og anatomisk (gsch .ind et al., 2012)., Da der imidlertid ikke er nogen aktuelle data tilgængelige vedrørende LO ‘ s rolle i dette system, forbandt vi dem med de to andre regioner på flere plausible måder.
den første tilfældige BMS viste, at LO i winnerinner-modelfamilien er sammenkoblet med både FFA og OFA, og forbindelserne er tovejs. Derfor fremhæver det, at LO kan have en direkte strukturel forbindelse til FFA., Med modellering alle mulige modulerende virkninger fandt vi, at en sub-familie af modeller, der vandt, hvor både objekt og ansigt indgange ind i systemet via LO, lad os antage, at LO spiller en generel og vigtig input region rolle. Siden tidligere funktionelle forbindelsesundersøgelser startede alle analysen af ansigtsbehandlingsnetværket på niveau med iog, svarende til OFA (Fairhall and Ishai, 2007; Ishai, 2008; Cohen Kadosh et al., 2011; Dima et al., 2011; Foley et al., 2012) det er ikke overraskende, at de overså LO ‘ s betydelige rolle., Imidlertid er ansigter faktisk en særskilt kategori af visuelle objekter, hvilket antyder, at neuroner, der er følsomme over for objekter og former, også skal aktiveres, i det mindste til en vis grad, af ansigter. Faktisk antyder enkeltcelleundersøgelser af ikke-humane primater, at den underordnede temporale Corte., den foreslåede homolog af human LO i makakhjernen (Denys et al., 2004; Sa .amura et al., 2006) har også neuroner, der reagerer på ansigter (Perrett et al., 1982, 1985; Desimone et al., 1984; Hasselmo et al., 1989; Young and Yamane, 1992; Sugase et al., 1999)., Den intime forbindelse af LO til OFA og FFA, foreslået af den nuværende undersøgelse, kunne understrege det faktum, at ansigter og genstande ikke behandles helt separat i den ventrale visuelle vej, en konklusion understøttet af nylige funktionelle billeddannelsesdata også (Rossion et al., 2012). Den modulerende effekt af ansigt input på LO–FFA forbindelse foreslår, at LO skal spille en rolle i ansigt-behandling, sandsynligvis knyttet til tidligere, strukturelle behandling af ansigter, en opgave der tidligere er sat ned, for det meste til OFA (Rotshtein et al., 2005; fo Fox et al. , 2009)., Endelig gruppe-niveau parametre af vinderen model viser, at forbindelsen mellem LO, OFA, og FFA er primært hierarkisk, og den feedback forbindelser fra FFA mod OFA og LO spiller en svagere rolle i systemet, i det mindste under anvendt fiksering opgave.
afslutningsvis foreslår vi ved at modellere den effektive forbindelse mellem ansigtsrelevante områder og LO, at LO spiller en betydelig rolle i behandlingen af ansigter.,
Erklæring om interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
dette arbejde blev støttet af Deutsche Forschungsgemeinschaft (KO 3918/1-1) og University of Regensburg. Vi vil gerne takke Ingo Keck og Michael Schmitgen for diskussion.
Campbell, R. (2011). Speechreading og Bruce-Young model af ansigtsgenkendelse: tidlige resultater og den seneste udvikling. Br. J. Psychol. 102, 704–710.,
Pubmed Abstract | Pubmed Fuld Tekst | CrossRef Fuld Tekst
Fairhall, S. L., og Ishai, A. (2007). Effektiv forbindelse inden for det distribuerede kortikale netværk til ansigtsopfattelse. Cereb. Corte.17, 2400-2406.
Pubmed Abstract | Pubmed Fuld Tekst | CrossRef Fuld Tekst
Hoffmann, E. A., og Haxby, J. V. (2000). Distinkte repræsentationer af blik og identitet i det distribuerede menneskelige neurale system til ansigtsopfattelse. Nat. Neurosci. 3, 80–84.
PubMed Abstract | PubMed Full te Abstractt/CrossRef Full Te.t
Kourt ,i ,.. ,, og kan Kanisher, N. (2001). Repræsentation af opfattet objektform af det menneskelige laterale occipitale kompleks. Videnskab 293, 1506-1509.
Pubmed Abstract | Pubmed Fuld Tekst | CrossRef Fuld Tekst
MacKay, D. J. C. (2003). Information teori, følgeslutning, og læring algoritmer. Cambridge: Cambridge University Press.
Nasanen, R. (1999). Rumlig frekvensbåndbredde, der anvendes til genkendelse af ansigtsbilleder. Vision Res. 39, 3824-3833.
Pubmed Abstract | Pubmed Fuld Tekst | CrossRef Fuld Tekst
Rossion, B., Schiltz, C., og Crommelinck, M., (2003b). De funktionelt definerede højre occipitale og fusiform “ansigtsområder” diskriminerer roman fra visuelt kendte ansigter. Neuroimage 19, 877-883.
CrossRef Fuld tekst
Sa .amura, H., Orban, G. A., and Vogels, R. (2006). Selektivitet af neuronal tilpasning svarer ikke til responsselektivitet: en enkeltcelleundersøgelse af fMRI-tilpasningsparadigmet. Neuron 49, 307-318.
PubMed abstrakt / PubMed Fuld tekst / CrossRef Fuld tekst
















Skriv et svar