Indledning
Kloroplaster repræsenterer en strukturel funktion af plante celler og støtte plante overlevelse via deres primære metabolisme og høj-niveau funktioner (Kirk og Tilney-Bassett, 1978; Mulle, 1988; López-Juez og Pyke, 2005). Under plantevegetativ vækst indeholder bladceller en meget homogen population af chloroplaster med hensyn til størrelse og form., Antallet af grønkornene per celle er opnået ved binær fission af præ-eksisterende organeller og opdeling i to datterceller under celledeling (Birky, 1983; Possingham og Lawrence, 1983). Således er regulering af chloroplastnummeret i en given celletype afgørende for den cellulære funktion og genetiske arv af chloroplaster.
for at undersøge arten af bestemmelse af chloroplastnummer i planteceller har bladmesofylceller af repræsentative arter spillet en vigtig rolle ., Disse celler er fysiologisk vigtige for fotosyntese og viser en høj grad af strukturel og funktionel homogenitet. Tidlig systematisk observation analyser af isolerede væv og celler (Boasson og Laetsch, 1969; Possingham og Saurer, 1969; Boffey et al., 1979; Lamppa et al.,, 1980; Thomas og Steg, 1983; Pyke og Agterliget, 1991) givet mange nyttige oplysninger om kloroplast antal bestemmelse, herunder forestillingen om, at grønkornene (plastider) er ikke syntetiseret de novo men formere ved deling og observation, at bladet mesophyll kloroplast tal er følsom over for forskellige miljø-og plante-endogene faktorer. Med hensyn til sidstnævnte har lyset i spinat en positiv indvirkning på chloroplastinddeling under bladskivekultur sammenlignet med svage eller mørke forhold (Possingham og La .rence, 1983)., I de første blade af hvede er cellevolumen positivt korreleret med chloroplastproliferation (Ellis og Leech, 1985; Pyke og Leech, 1987). I Arabidopsis, den genetiske baggrund påvirker kloroplast spredning; den gennemsnitlige kloroplast antal pr celle i første blade er 121 i Landsberg erecta (Ler) ecotype og 83 i Wassilewskija (Ws) ecotype (Pyke og Agterliget, 1994; Pyke et al., 1994)., Blad mesophyll celler, også har bidraget til forståelsen af de genetiske kontrol af kloroplast division; for eksempel screening mutanter nedsat i kloroplast spredning og karakterisering af genet funktioner, der er involveret i kloroplast division har afsløret over 20 gener, der koder for kloroplast division maskiner, komponenter eller grønkorn-regulatoriske faktorer (Gao og Gao, 2011; Miyagishima et al., 2011; Basak and Møller, 2013; Osteryoung and Pyke, 2014; Li et al., 2017).
derimod er undersøgelser af replikation af chloroplaster i ikke-mesofylceller (f. eks.,, fortovsceller i bladepidermis; Itoh et al., 2018) er knappe. For nylig er reguleringen af chloroplastinddeling rapporteret at variere mellem bladvæv (Fuji .ara et al., 2018; Itoh et al., 2018), selvom den detaljerede mekanisme forbliver ukendt. Derudover, mens analyser af suspension er dannet VED-2-celler og blade mesophyll protoplasts i tobak og skyde apikale meristem og blade oprindelige celler i Arabidopsis (Nebenführ et al., 2000; Sheahan et al.,, 2004; Seguí-Simarro og Staehelin, 2009), har givet en større indsigt, hvordan grønkorn (plastid) partitionering er reguleret i planter, er stadig uklart. På trods af en betydelig indsats forbliver der således grundlæggende spørgsmål i chloroplastforskning, såsom (i) hvordan koordineres chloroplastnummer pr.celle i plantevæv og (ii) hvordan reguleres chloroplastpartitionering ved celledeling.,
Historie af Forskning på Vagt Celle Kloroplast-Nummer
Stomatale GCs i skyde epidermis generelt indeholder kloroplaster og kontrol gas udveksling mellem blad mesophyll og atmosfæren (Sachs, 1875; Taiz et al., 2015; se figur 1A). Den første undersøgelse af GC chloroplast nummer i blade blev udført over et århundrede siden i naturligt dyrkede Drosera planter (Macfarlane, 1898)., Denne undersøgelse viste, at de ligesom andre plante-og cell strukturelle egenskaber, GC kloroplast antal pr celle i en formodede hybrid stammer fra en krydsning mellem Drosera filiformis og Drosera intermedia var mellemliggende mellem de to arter, hvilket betyder, at GC kloroplast-nummer kan anvendes til at bestemme den genetiske sammensætning af en plante. Vigtige bemærkninger blev efterfølgende rapporteret om forskellene i GC kloroplast blandt plantearter (Sakisaka, 1929), og den relativt stabile kloroplast nummer i GCs i bladet epidermis af morbær (Morus spp.,; Hamada og Baba, 1930) og i modne blade af flere Brassica-arter (Iura, 1934). Endvidere afslørede analyse af autopolyploid sukkerroer (Beta vulgaris) planter, at GC-chloroplastnummeret i blade er positivt korreleret med planternes nukleare ploidiniveau (Mochi .uki og Sueoka, 1955). Mere dybtgående og omfattende analyser blev derefter udført ved hjælp af forskellige planteprøver for at undersøge forholdet mellem chloroplastnummer og stomatal størrelse med ploidiniveauet (f.eks. Frandsen, 1968)., I disse analyser, kloroplast tælle på stomatale (GC par) niveau var ofte har vedtaget, og som er udelukket effekten af forudindtaget kloroplast fordeling mellem parret GCs (fx, Mochizuki og Sueoka, 1955; Frandsen, 1968), der afslører, at den gennemsnitlige GC kloroplast-nummer, i blade eller kimbladene i omkring 80 arter, varianter, eller hybrider varierede fra 2,8 til 40.0 i diploider (2×), og 5.0 til 73,5 i tetraploids (4×). Hertil kommer, hel-genom dobbeltarbejde begivenheder i planter (dvs., 1 to til 2., 2. til 4., etc.) forårsagede en ca.1.,7 gange stigning i GC-chloroplastantal med høj tro (gennemgået i Butterfass, 1973). Disse resultater opfordres undersøgelser ploidy niveau i de forskellige væv og planter opnået via vævskultur, krydser, eller naturlig dyrkning i kombination med kemisk (f.eks colchicin) eller strålebehandling, behandlinger (fx Jacobs og Yoder, 1989; Singsit og Veilleux, 1991; Qin og Rotino, 1995)., Mens GC kloroplast-nummer er blevet undersøgt i stomatale biologi (Lawson, 2009) og cytologi at forstå kloroplast multiplikation (Butterfass, 1979; se nedenfor), er det i vid udstrækning har fungeret som en pålidelig og bekvem markør til påvisning af hybrider, arter og varianter, og til estimering af ploidy niveauer af mål plantevæv.
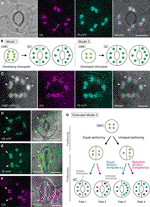
Figur 1 Forenklede modeller og mikroskopi dokumentation for kontrol af kloroplast-nummer i stomatale vagt celler (GCs)., (A) En typisk stomi (GC par) i abaxial epidermis af Arabidopsis bladpladen at udtrykke en stomi-målrettet fusion af transit peptid (TP) med cyan fluorescerende protein (FFP; TP-CFP). B) to modeller for bestemmelse af chloroplasttal i GCs, der involverer enten chloroplastpartitionering (model 1) eller både chloroplastproliferation og partitionering (model 2) under GC-udvikling fra guard-moderceller (GMC ‘ er). C) et GC-par i adaksial epidermis af Arabidopsis-bladblad, der udtrykker TP-CFP og FTS .1 smeltet sammen med det grønne fluorescerende protein (GFP; FTS .1-GFP)., (D–F) GC par i abaxial epidermis af Arabidopsis blad klinge med (D, E) eller uden (F) udtrykket af TP smeltet til gul fluorescerende protein (YFP; TP-YFP). (F) Chlorophyll autofluorescens (Chl) blev anvendt som kloroplastmarkør., (G) Udvidet model 2, svarende til inddragelse af lige og ulige kloroplast partitionering følgende GMC division og efterfølgende opdeling af GC kloroplaster med lige (blå linje) eller selektivt (rød linje) division kompetence, hvilket vil resultere i fire typer af kloroplast antal bestemmelse (Gruppe 1-4) i slutningen af stomatale udvikling af Arabidopsis blade. (A, C–f) Epifluorescensmikroskopi blev udført med et Olympus i .71 inverteret mikroskop ved anvendelse af plantematerialer som tidligere beskrevet (Fuji .ara et al., 2017, Fuji .ara et al., 2018)., Fluorescens signaler af klorofyl, CFP, GFP, og YFP er pseudo-farvet i magenta, cyan, grøn (kun i fusionerede billede), og grøn, henholdsvis. Indikationer i paneler er som følger: pilespids, FTS .1-ringen; pil, forstørret GC-chloroplast; stiplet linje, celleform. Vægtstang = 10 µm.
Nytte af Blade Vagt Celler Til Analyse Af Kloroplast Antallet Kontrol
Blad mesophyll celler har længe været ansat som primær model til analyse af kloroplast-nummer., Mens de har fordele for undersøgelse af virkningerne af miljømæssige forhold på kloroplast division (fx, lys, cytokinin signaler; Boasson og Laetsch, 1969; Possingham og Lawrence, 1983; Okazaki et al., 2009; Chiang et al. , 2012), er de begrænset i nogle henseender. For det første varierer blad mesophyll celler i størrelse og form og fordeles dybt inde i bladet, hvilket gør det vanskeligt at manipulere intakte væv., For det andet kan følsomheden af blad mesophyll chloroplastproliferation for miljøbelastning og plantevækstbetingelser forhindre pålidelige sammenligninger mellem undersøgelser. Bladet mesophyll kloroplast antal pr celle i Arabidopsis Columbia (Col) ecotype er blevet rapporteret forskelligt, som 76 (Kinsman og Pyke, 1998), 80-100 (Stokes et al., 2000), 70 (Tirlapur og König, 2001), 41 (Yoder et al., 2007), 40-60 (Okaakiaki et al ., 2009), og 30-40 (ka kaade et al ., 2013)., Desuden er det næsten umuligt at vurdere bidraget fra grønkornenes partitionering til den endelige kloroplast antal pr mesophyll celle under blad udvikling, selv om dette menes at være bestemt af balancen mellem hastigheden af celledeling og sats af kloroplast division. For at afdække mekanismen for chloroplast nummerkontrol i vegetative bladceller kræves et modelsystem, der overvinder ovenstående problemer.
Stomatal GCs (se figur 1A) udviser egenskaberne ved et modelsystem til forståelse af mekanismen for chloroplast nummerkontrol., GCs er meget ensartede i størrelse og form inden for et væv, og deres spredte, men tætte fordeling i det yderste lag af skud letter deres påvisning ved lys-og fluorescensmikroskopi. GCs er også afledt af protodermal celler i skyde apikale meristem eller fra embryonale epidermale celler, og deres udviklings-sekvens gennem meristemoids (en stomatale forløber med meristematic aktivitet) og vagt mor celler (GMCs; en forløber for GC par) er etableret i detaljer (Zhao og Sæk, 1999; Nadeau og Sæk, 2002; Kalve et al., 2014)., Sent stomatale udvikling indebærer en enkelt runde af symmetrisk GMC division, som muliggør vurdering af kloroplast-distribution og-partitionering før og efter cytokinese. Set fra praktiske eksperimenter er bladgcs egnede til mikroskopi. Det blev tidligere vist, at kloroplast antal pr celle i bladet GCs af Sinapis alba var mindre påvirket af forskellige lysforhold end i blad mesophyll celler (Wild og Wolf, 1980)., Derudover er forskellen i GC-chloroplastnummer i bladblader relativt lille blandt de tre Arabidopsis-økotyper Col, Ler og .s (Fuji .ara et al ., 2018). Desuden endoreduplikation, som påvirker udviklingen af blade mesophyll, fortovet, og trichome celler, er ikke blevet registreret i Arabidopsis blad GCs (Melaragno et al., 1993), hvilket ville sikre fortolkningerne af chloroplast-taldata på 2C-niveauet af celler., Tilsammen antyder disse rapporter, at bladgcs potentielt er en fremragende model til systematisk analyse af chloroplastnummerdynamik i en bestemt cellelinje.
Arabidopsis Blad Vagt Celler som en Model til at Studere Kontrol af Kloroplast-Nummer
I historien om GC kloroplast forskning, kloroplast tælle på stomatale (GC par) plan har tjent en lige så vigtig rolle i fastsættelsen af kloroplast-nummer som at tælle på de enkelte GC-niveau. Begge metoder producerer det samme gennemsnitlige kloroplasttal (Butterfass, 1973)., Når variationen i chloroplastfordeling i parret GCs og dens underliggende mekanisme er genstand for fokus, er detaljerede oplysninger om chloroplaster på det individuelle celleniveau, dvs.deres størrelse, form og intracellulær lokalisering, afgørende. Chloroplast (plastid) proliferation under GMC–GC differentiering blev tidligere undersøgt i flere plantearter (Butterfass, 1973, Butterfass 1979)., Disse undersøgelser har foreslået to modeller til bestemmelse af terminalen kloroplast nummer i GCs i forskellige plante arter (Figur 1B): en (model 1; sukkerroer) omfatter kun kloroplast partitionering ved GMC division, og den anden involverer ikke kun kloroplast partitionering, men også grønkorn spredning i GC udvikling.
i en tid med molekylær genetik, genomik, celleafbildning og andre tværfaglige analyser er der mange muligheder for yderligere karakterisering af chloroplastpartitioneringsmekanismen., Arabidopsis leaf GCs kan være et af de bedste modelsystemer til dette formål. Flere undersøgelser har undersøgt GC-chloroplastnummeret i blade eller cotyledoner af Arabidopsis (Hoffmann, 1968; Pyke and Leech, 1994; Pyke et al., 1994; Robertson et al., 1995; Keech et al., 2007; Chen et al. , 2009; Yu et al., 2009; Higaki et al., 2012; Fuji 2012ara et al., 2018). Disse GCs udviser et beskedent antal chloroplaster, der spænder fra 3, 5 til 5, 5 i gennemsnit. Til dato har ingen undersøgelser undersøgt ændringerne i chloroplast (plastid) nummer under stomatal udvikling., Dog mikroskopiske beviser fra stomatal udvikling analyser (f 1999 Zhao og Sack, 1999; Hache.et al., 2011) og vores foreløbige observationer indikerer, at GMC ‘ er kan indeholde mindre antal udviklende chloroplaster end GCs, og at chloroplastproliferation kan forekomme under GC-differentiering. For at teste dette blev dannelsen af chloroplastdivisionsmaskineriet i GCS overvåget med sonden FTS .1 smeltet til det grønne fluorescerende protein (FTS .1-GFP) (Fuji .ara et al., 2008)., En transgene linje, samtidigt med at udtrykke en transit-peptid (TP)-smeltet fælles FISKERIPOLITIK og FtsZ1-GFP til at visualisere stroma og FtsZ1 ring, henholdsvis, blev gennemgået af epifluorescens-mikroskopi (Fujiwara et al., 2017). Ekspanderende bladblomstrer (femte blade af 4 uger gamle frøplanter) blev ansat. Som et resultat blev GCS med symmetrisk indsnævrende chloroplaster detekteret (figur 1C). Disse chloroplaster dannede FTS .1-ringen, en chloroplastdelingsring på den stromale overflade af den indre kuvertmembran i blad mesophyll og fortovsceller (Vitha et al., 2001; Fuji Fuara et al.,, 2008), på det ækvatoriale indsnævringssted. I overensstemmelse med stomatalmønsteret i Arabidopsis bladudvikling (Donnelly et al., 1999; Andriankaja et al., 2012) blev opdeling af chloroplaster påvist i det sene såvel som tidlige stadium af bladudvidelse. Således er model 2 sandsynligvis den bedste pasform til Arabidopsis leaf GCs.
sammenhæng mellem Chloroplastproliferation og-ekspansion i Arabidopsis-Bladbeskyttelsesceller
endvidere blev der observeret en uventet fænotype af GC-chloroplastmorfogenese i modne GCs (figur 1D, e)., Når epidermal skaller af fuldt udvidet blade (tredje–fjerde blad blade af 4-ugers-gamle planter) fra en TP-smeltet gul fluorescerende protein (YFP) line var mikroskopisk er karakteriseret (FL6-5 linje, Fujiwara et al., 2018) viste nogle stomata ulige chloroplastfordelingsmønstre i GC-par, mens de fleste bladstomata viste lige eller lignende chloroplastfordelingsmønstre (Robertson et al., 1995; Fuji 1995ara et al., 2018)., Inden for GC par af en stomi, størrelsen af chloroplaster i GC indeholdende mindre antal chloroplaster var større end i de andre GCs i parret indeholder større antal chloroplaster (tal 1D, E). På denne måde opretholder GCS sandsynligvis det samlede chloroplastvolumen pr. Forstørrede chloroplaster repræsenterede den terminale fænotype og kunne ikke længere opdele sig i udvidede blade. Disse resultater blev bekræftet i flere uafhængige eksperimenter, uanset ekspressionen af et TP-smeltet fluorescerende protein til stroma-mærkning (figur 1F).,
denne GC-chloroplastfænotype fortolkes som en kompensationsmekanisme for chloroplastudvidelse, hvilket var veldokumenteret i blad mesophyll celler defekt i kontrollen af chloroplast division (Pyke og Leech, 1994; Pyke et al., 1994). Til dato har kun én undersøgelse (Ellis og Agterliget, 1985) har rapporteret en negativ korrelation mellem kloroplast-nummer og kloroplast størrelse i bladet mesophyll celler af hvede, der henviser til, at mange undersøgelser har rapporteret en positiv sammenhæng mellem celle volumen og kloroplast nummer i normal blade mesophyll celler (Leech og Pyke, 1988; Pyke, 1997)., Mens ubalancer i GC-chloroplasttal forekommer ved lav frekvens (Fuji .ara et al ., 2018), kloroplast heterogenitet i GC par indikerer, at ulige kloroplast partitionering kan udløse differentieret kloroplast vækst mellem vildtype-bladet celler i Arabidopsis, på trods af symmetrisk celledeling.
chloroplastkompensationseffekten i GCs kan være mindre streng end i blad mesophyll celler., GCs kan være i stand til at modstå knaphed eller fuldstændigt tab af det samlede kloroplast volumen per celle i alvorlig nedsat kloroplast division mutanter, som i Arabidopsis arc6 og atminE1 og tomat suffulta, der henviser til, at mange mutant GCs viste nedsat kloroplast-nummer og udvidede kloroplast størrelse på samme måde som bladet mesophyll celler (Robertson et al., 1995; Forth and Pyke, 2006; Chen et al., 2009; Fuji Fuara et al., 2018)., I en sen chloroplastdivisionsmutant, arc5, var reduktionen i GC-chloroplastantal ikke forbundet med en signifikant stigning i chloroplaststørrelse, i modsætning til i blad mesophyll celler (Pyke og Leech, 1994). En lavere grad af chloroplastudvidelse i GCs end i mesophyll celler (Pyke and Leech, 1994; Barton et al., 2016), og variationen i kloroplastudvidelse blandt GCs, kan ligge til grund for et så bredt tilladt område af det samlede kloroplastvolumen pr., Endvidere kan timingen af chloroplastinddeling under GMC–GC-differentiering påvirke den terminale GC-chloroplastfænotype betydeligt. Selvom der kræves yderligere detaljeret karakterisering for at løse dette problem, forekommer det sandsynligt, at Arabidopsis leaf GCs repræsenterer et system til at undersøge de uudnyttede aspekter af chloroplast nummerkontrol i planteceller.,
En fungerende Model For Kloroplast Antal Bestemmelse i Arabidopsis Blad Vagt Celler
På baggrund af ovenstående, foreslår vi en fungerende model (en udvidet model 2) til analyse af kloroplast-nummer i GCs (Figur 1A). Det endelige chloroplasttal pr. GC bestemmes ved chloroplastpartitionering ved GMC-division og chloroplastproliferation i GCs. Under GMC-division kan chloroplaster gennemgå enten lige eller ulige partitionering. Under chloroplastproliferation vil GC-chloroplaster spredes med enten lige (blå linje) eller selektiv (magenta linje) divisionskompetence., For eksempel, hvis lige partitionerede chloroplaster besidder ækvivalent divisionskompetence, vil der forekomme lige chloroplastnumre i GC-parret (Fate 1). Hvis ulige partitionerede chloroplaster besidder ækvivalent divisionskompetence, vil chloroplaster stige med samme hastighed inden for GC-parret (Fate 3). Hvis selektiv chloroplast division forekommer i GCs, vil balancen af chloroplast nummer i GC parret ændre sig efter GMC division (Fates 2 og 4)., Det er i øjeblikket svært at finde støtte til “selektiv chloroplast division”, men hvis skæbner 1 og 4 faktisk dominerer i GCs, så kan de have en mekanisme, der styrer det samlede chloroplastvolumen pr. Modellen rejser to spørgsmål: (I) er GC-chloroplaster korrekt opdelt i datterceller, og hvordan partitionerer de? Og ii) reguleres GC-chloroplasters kompetencefordeling koordinerende?,
med Hensyn til spørgsmålet (i), om kloroplast arv sker ved tilfældig fordeling af flere kloroplaster i cytoplasmaet eller ved positiv kloroplast partitionering mekanisme(r) har længe været en bekymring (Butterfass, 1969; Birky, 1983; Hennis og Birky, 1984; Nebenführ, 2007; Sheahan et al., 2016). Interessant nok har Arabidopsis arc6, blad-eller cotyledon-GCs nul til tre chloroplaster, og i chloroplast-mangelfulde GCs findes der stadig ikke-fotosyntetiske plastider i vesikulær til langstrakt form (Robertson et al., 1995; Chen et al., 2009; Fuji Fuara et al., 2018)., Ingen GCs blottet for plastider per se er blevet fundet i arc6, og ingen forklaring på dette har været tilfældet, på trods af afbrydelsen af kloroplast division apparater (Vitha et al., 2003). Det vil derfor være vigtigt at undersøge replikationen og morfologien af arc6-chloroplaster i stomatal afstamningsundersøgelser. Arabidopsis mutant forskning kan også give en anden anelse om dette problem., Observationen af, at 18% af cotyledon GCs i den krøllede blad (crl) – mutant ikke indeholder nogen plastidiske strukturer i cytoplasmaet, mens 100% af blad mesophyll-cellerne indeholder en til fire forstørrede chloroplaster (Asano et al., 2004; Chen et al., 2009), er af stor betydning. CRL er et kloroplast ydre kuvert protein med en ukendt funktion. Forståelse af CRL-proteinfunktion kan give indsigt i mekanismen / mekanismerne for chloroplastpartitionering., Analyse af kloroplast spredning og fordeling i bladet mesophyll celler i Arabidopsis arc mutanter og andre transgene linier har støttet forskning i spredning og fordeling af ikke-mesophyll plastider. Ligeledes kan resultater opnået i GCs overføres til andre cellesystemer.
endelig bemærkning
GC-modellen åbner mange udsigter til udvikling af chloroplastbiologi. For eksempel, mens cytoskeletale systemer er kendt for at regulere chloroplastmorfologi, bevægelse og partitionering (Sheahan et al.,, 2016 ;adaada, 2016; Erickson og Schattat, 2018), har hvert regulatorisk gens rolle i chloroplastproliferation og opdeling i planter fået lidt opmærksomhed. På den anden side, når det bliver muligt at forringe GC kloroplast nummer eller morfologi via forskellige eksperimentelle strategier, ny indsigt i den molekylære kontrol af kloroplast morfogenese i stomatale slægt celler kan angives. Derudover, i forbindelse med kvantitative analyser af chloroplast nummer under stomatal udvikling, matematisk modellering kan tilbyde en ny vej til disse undersøgelser., Dette papir præsenterer aktuel viden om, hvordan GC-chloroplastnummer styres og fremhæver den potentielle anvendelighed af Arabidopsis leaf GCs til forståelse af chloroplastproliferation og partitionering.
datatilgængelighed Erklæring
alle datasæt for denne undersøgelse er inkluderet i artiklen / supplerende materiale.
forfatter Bidrag
MF udtænkt undersøgelsen og skrev manuskriptet. MF og som udført forsøgene. AS og RI gennemførte analyserne. Alle forfattere læste og godkendte det endelige manuskript.,
Finansiering
Dette arbejde blev støttet af Ministeriet for Uddannelse, Kultur, Videnskab og Teknologi Japan under KAKENHI (bevilling nr. 19K05831 at MF og 18K06314 at RI).
interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
forfatterne takker Dr. Nobuyuki kan .a .a (Sophia University) for rådgivning om arbejdet.
Birky, C. W., Jr., (1983)., Opdeling af cytoplasmatiske organeller ved celledeling. Int. Rev. Cytol. 15, 49-86.
Google Scholar
smør churn, T. (1969). Plastidfordelingen i mitose af de afsluttende cellemoderceller af haploid s .edenklee (Trifolium hybridum L.). Planta 84, 230-234. doi: 10.1007/BF00388108
PubMed Abstract | CrossRef Fuld Tekst | Google Scholar
smør churn, T. (1973). Kontrol af plastid division ved hjælp af nuklear DNA mængde. Protoplasma 76, 167-195. doi: 10.,1007/BF01280696
CrossRef Full Text | Google Scholar
Qin, X., Rotino, G. L. (1995). Chloroplast number in guard cells as ploidy indicator of in vitro-grown androgenic pepper plantlets. Plant Cell Tissue Organ Cult. 41, 145–149. doi: 10.1007/BF00051583
CrossRef Full Text | Google Scholar
Sachs, J. (1875). Textbook of Botany. Oxford: Clarendon Press.
Google Scholar
Thomas, M. R., Rose, R. J. (1983)., Plastid antal og plastid strukturelle ændringer i forbindelse med tobak mesophyll protoplast Kultur og plante regenerering. Planta 158, 329-338. doi: 10.1007/BF00397335
PubMed Abstract | CrossRef Fuld Tekst | Google Scholar
Tirlapur, U. K., König, K. (2001). Femtosekund nær-infrarøde lasere som et nyt værktøj til ikke-invasiv realtid høj opløsning time-lapse billeddannelse af chloroplast division i levende bundt kappe celler af Arabidopsis. Planta 214, 1-10. doi: 10.,1007/s004250100597
PubMed Abstract | CrossRef Fuld Tekst | Google Scholar
Wild, A., Ulv, G. (1980). Effekten af forskellige lysstyrker på hyppigheden og størrelsen af stomata, størrelsen af celler, antal, størrelse og klorofyl indhold af grønkornene i mesophyll og vagten celler i ontogeni af primær blade af Sinapis alba. `.. Pflan Penphysiol. Bd. 97, 325–342. doi: 10.1016/S0044-328X(80)80006-7
CrossRef Fuld Tekst | Google Scholar
















Skriv et svar