Introducere
procesarea neurală a fețelor este un subiect larg cercetat al științei cognitive. Pe baza studiilor imagistice funcționale, a înregistrărilor cu o singură celulă și a cercetărilor neuropsihologice, s-a sugerat că prelucrarea feței este efectuată de o rețea distribuită, care implică mai multe zone corticale ale creierului mamifer (Haxby et al., 2000; Marotta și colab., 2001; Rossion și colab., 2003a; Avidan și colab., 2005; Sorger și colab., 2007)., În timp ce amploarea acestei rețele de procesare a feței este în prezent în dezbatere intensă (Ishai, 2008; Wiggett și Downing, 2008), majoritatea cercetătorilor sunt de acord că există numeroase zone corticale activate de stimulii feței. Cel mai influent model de percepție a feței, bazat pe modelul original al lui Bruce și Young, 1986; Young și Bruce, 2011) propune o distincție între reprezentarea aspectelor invariante și variante ale percepției feței într-o manieră relativ independentă, separate într-un „nucleu” și o parte „extinsă” (Haxby et al., 2000)., Cele mai importante regiuni ale „rețelei de bază” sunt zone ale occipitalului și gyri fusiform lateral (FG). Zonele acestor două regiuni anatomice par a fi specializate pentru sarcini distincte: în timp ce zona occipitală a feței (OFA), situată pe girusul occipital inferior (IOG) pare să fie implicată în prelucrarea structurală a fețelor, zona fusiformă a feței (FFA) procesează fețele într-o manieră superioară, contribuind, de exemplu, la prelucrarea identității (Sergent et al., 1992; George și colab., 1999; Ishai și colab., 1999; Hoffman și Haxby, 2000; Rossion și colab., 2003a, b; Rotshtein și colab.,, 2005). În plus, aspectele schimbătoare ale fețelor (cum ar fi expresiile faciale, direcția privirii ochilor, expresia, mișcările buzelor (Perrett et al., 1985, 1990) sau citirea buzelor (Campbell, 2011) par a fi procesate în sulcusul temporal superior (STS; Puce et al., 1998; Hoffman și Haxby, 2000; Winston și colab., 2004). Cele trei domenii menționate mai sus (FFA, OFA, STS) formează așa-numitul „nucleu” al sistemului perceptual de prelucrare a feței (Haxby et al., 2000; Ishai și colab., 2005)., În timp ce informațiile de bază despre fețe sunt procesate de acest sistem de bază, informațiile complexe despre starea de spirit, nivelul de interes, atractivitatea sau direcția atenției celorlalți adaugă, de asemenea, informații pentru percepția feței și sunt procesate de un sistem suplimentar, așa-numitul „extins” (Haxby et al., 2000). Acest sistem conține regiuni ale creierului cu o mare varietate de funcții cognitive legate de prelucrarea aspectelor faciale schimbătoare (Haxby et al., 2000; Ishai și colab., 2005) și includ zone precum amigdala, insula, girusul frontal inferior, precum și cortexul orbitofrontal (Haxby et al.,, 2000, 2002; Fairhall și Ishai, 2007; Ishai, 2008).interacțiunile ariilor menționate mai sus sunt modelate în studiul de față folosind metode care calculează conectivitatea eficientă între ariile corticale. Modelarea cauzală dinamică (DCM) este o metodă utilizată pe scară largă pentru a explora conectivitatea eficientă între regiunile creierului. Este o abordare generică pentru modelarea influenței reciproce a diferitelor zone ale creierului una asupra celeilalte, bazată pe activitatea fMRI (Friston et al., 2003; Stephan și colab., 2010) și estimarea modelului de inter-conectare a ariilor corticale., DCM-urile sunt modele generative de răspunsuri neuronale, care oferă estimări „a posteriori” ale conexiunilor sinaptice în rândul populațiilor neuronale (Friston et al., 2003, 2007). Existența rețea distribuită de procesare a feței a fost confirmată de conectivitate funcțională analiza de Ishai (2008), care a susținut că nodul central de procesare a feței este laterale FG, conectat la ordin inferior zonele de JURNAL, precum și pentru a STS, amigdala, și zonele frontale. Prima încercare de a dezvălui rețeaua de procesare a feței în cazul expresiilor faciale dinamice realiste a fost făcută de Foley et al. (2012)., Aceștia au confirmat rolul zonelor IOG, STS și FG și au constatat că rezistența conexiunii dintre membrii rețelei centrale (OFA și STS) și ai sistemului extins (amigdala) este crescută pentru procesarea gesturilor încărcate de afectare. În prezent, mai multe studii au elaborat înțelegerea noastră asupra rețelei de procesare a feței și au relevat o legătură directă între amigdală și FFA și rolul acestei conexiuni în percepția fețelor înfricoșătoare (Morris et al., 1996; Marco și colab., 2006; Herrington și colab., 2011)., În cele din urmă, efectul funcțiilor cognitive superioare asupra percepției feței a fost, de asemenea, modelat prin testarea conexiunilor cortexului orbitofrontal la rețeaua centrală (Li et al., 2010). A fost găsit (Li et al., 2010) că cortexul orbitofrontal are un efect asupra OFA, care modulează în continuare procesarea informațiilor FFA.în timp ce studiile anterioare de conectivitate eficiente au dezvăluit detaliile rețelei de procesare a feței legate de diverse aspecte ale percepției feței, au ignorat simplul fapt că fețele pot fi considerate și obiecte vizuale., Știm dintr-un corp mare de experimente care obiecte vizuale sunt procesate de către o distribuite corticale de rețea, inclusiv devreme ariile vizuale, occipito-temporal, și ventral–temporale cortices, în mare parte se suprapun cu fata-rețea de prelucrare (Haxby et al., 1999, 2000, 2002; Kourtzi și Kanwisher, 2001; Ishai și colab., 2005; Gobbini și Haxby, 2006, 2007; Haxby, 2006; Ishai, 2008). Una dintre zonele majore de prelucrare a obiectelor vizuale este cortexul occipital lateral (LOC), care poate fi împărțit în două părți: partea anterioară–ventrală (PF/LOa) și partea caudală–dorsală (LO; Grill-Spector et al.,, 1999; Halgren și colab., 1999). LOC a fost descris pentru prima dată de Malach et al. (1995), care a măsurat o activitate crescută pentru obiecte, inclusiv fețe celebre, în comparație cu obiectele amestecate (Malach et al., 1995; Grill-Spector et al., 1998a). De atunci, occipitalul lateral (LO) este considerat în primul rând ca o zonă selectivă a obiectului, care totuși se dovedește invariabil că are o activare ridicată și pentru fețe (Malach et al., 1995; Puce și colab., 1995; Lerner și colab., 2001), în special pentru cele inversate (Aguirre et al., 1999; Haxby și colab., 1999; Epstein și colab.,, 2005; Yovel și Kanwisher, 2005).astfel ,este destul de surprinzător faptul că, în timp ce mai multe studii s-au ocupat de conectivitatea eficientă a zonelor de procesare a feței, niciunul dintre ele nu a considerat rolul LO în rețea. Într-un studiu fMRI anterior am constatat că LO are un rol crucial în competiția senzorială pentru stimulii feței (Nagy et al., 2011)., Activitatea de LO a fost redus cu prezentarea simultan prezentat concurente stimuli și acest răspuns reducere, care reflectă senzoriale concurența între stimuli, a fost mai mare atunci când cei din jur stimul a fost o fata, comparativ cu o Fourier-faza studiu randomizat de zgomot de imagine. Acest rezultat a susținut, de asemenea, ideea că LO poate juca un rol specific în percepția feței. Prin urmare, aici am explorat în mod explicit, folosind metode de conectivitate eficientă, modul în care LO este legat de FFA și OFA, membri ai rețelei centrale propuse de percepție a feței (Haxby et al., 1999, 2000, 2002; Ishai și colab., 2005).,
materiale și metode
subiecți
douăzeci și cinci de participanți sănătoși au luat parte la experiment (11 femele, mediană: 23 ani, min.: 19 ani, max.:35 ani). Toți au avut o viziune normală sau corectată la o viziune normală (auto-raportată), niciunul dintre ei nu a avut boli neurologice sau psihologice. Subiecții și-au furnizat consimțământul informat în scris în conformitate cu protocoalele aprobate de Comitetul etic al Universității din Regensburg.,
Stimuli
Subiectele au fost de nivel central, prezentat de gri-scară se confruntă, non-sens obiecte, și Fourier randomizat versiuni ale acestor stimuli, creat de un algoritm (Nasanen, 1999), care înlocuiește faza spectrului de frecvențe cu valori aleatoare (variind de la 0° la 360°), lăsând amplitudinea spectrului de imaginea intactă, în timp ce eliminarea orice formă de informații. Fețele erau imagini digitale full-front cu 20 de bărbați tineri și 20 de femei tinere. Acestea au fost potrivite în spatele unei măști de formă rotundă (3,5° diametru) eliminând contururile exterioare ale fețelor (a se vedea o imagine de probă din Figura 1)., Obiectele erau non-sens, obiecte redate (n = 40) având aceeași dimensiune medie ca și masca de față. Luminanța și contrastul (adică abaterea standard a distribuției luminanței) stimulilor au fost echivalate prin potrivirea histogramelor de luminanță (luminanța medie: 18 cd/m2) folosind Photoshop. Stimulii au fost proiectați înapoi prin intermediul unui proiector video LCD (JVC, DLA-G20, Yokohama, Japonia, 72 Hz, rezoluție 800 × 600) pe un ecran circular translucid (app. 30 ° Diametru), plasat în interiorul găurii scanerului la 63 cm de observator., Prezentarea stimulului a fost controlată prin intermediul software-ului e-prime (Psychological Software Tools, Pittsburgh, PA, SUA). Fețele, obiectele și imaginile de zgomot Fourier au fost prezentate în blocuri ulterioare de 20 s, intercalate cu 20 s de perioade goale (fundal gri uniform cu o luminanță de 18 cd/m2). Stimulii au fost prezentați pentru 300 ms și au fost urmați de un ISI de 200 ms (2 Hz) într-o ordine aleatorie. Fiecare bloc a fost repetat de cinci ori. Participanților li sa cerut să se concentreze continuu pe o marcă de fixare prezentată Central., Aceste runde localizatoare funcționale au făcut parte din alte două experimente de percepție a feței, publicate în altă parte (Nagy et al., 2009, 2011).

Figura 1. Exemple de stimuli ai experimentului. Toate imaginile au fost la scară gri, aceeași dimensiune, luminanță și contrast. Panoul din stânga prezintă o față, caracteristici specifice genului (cum ar fi părul, bijuteriile etc.) a fost ascunsă în spatele unei măști ovale. Panoul din mijloc prezintă un eșantion obiect geometric non-sens, în timp ce panoul din dreapta arată versiunea Fourier-fază randomizat de obiecte, utilizate ca stimuli de control.,
achiziția și analiza datelor
imagistica a fost efectuată utilizând un scaner de cap MR 3-T (Siemens Allegra, Erlangen, Germania). Pentru seria funcțională am achiziționat continuu imagini (29 de felii, înclinate la 10° față de axial, T2 * secvență EPI ponderată, TR = 2000 ms; TE = 30 ms; Unghi flip = 90°; matrice 64 × 64; rezoluție în plan: 3 mm × 3 mm; grosime felie: 3 mm). Imaginile sagitale de înaltă rezoluție T1 ponderate au fost achiziționate folosind o secvență EPI de magnetizare (MP-RAGE; TR = 2250 ms; TE = 2.,6 ms; 1 mm dimensiune izotropă voxel) pentru a obține o Scanare structurală 3D (pentru detalii, vezi Nagy et al., 2011).
Funcțional imaginile au fost corectate pentru achiziționarea de întârziere, realiniat, normalizat la MNI spațiu, resampled la 2 mm x 2 mm x 2 mm rezoluție și spațial netezite cu un nucleu Gaussian de 8 mm FWHM (SPM8, bun venit la Departamentul de Imagistica Neuroscience, Londra, marea BRITANIE; pentru detalii, a analizei datelor, a se vedea Nagy et al., 2011).
selecția VOI
În primul rând, au fost selectate volume de interese (VOI), pe baza constrângerilor de activitate și anatomice (inclusiv mascarea regiunilor cerebrale relevante)., Zonele selective ale feței au fost definite ca o zonă care prezintă o activare mai mare pentru fețe în comparație cu imaginile și obiectele cu zgomot Fourier. FFA a fost definit în girusul Fusiform Lateral, în timp ce OFA în IOG. LO a fost definit din obiectul > Contrastul zgomotului Fourier și al imaginilor feței, în girusul Occipital Mijlociu. Selecția VOI s-a bazat pe contrastele T ajustate cu contrastul F, (p < 0.005 necorectate cu o dimensiune minimă a clusterului de 15 voxeli)., VOIs au fost sferice cu o rază de 4 mm în jurul activării vârfului (pentru coordonatele individuale, a se vedea tabelul A1 din anexă). Variația explicată de prima variație proprie a semnalelor îndrăznețe a fost peste 79%. Doar zonele din emisfera dreaptă au fost utilizate în analiza DCM actuală, deoarece mai multe studii indică rolul dominant al acestei emisfere în percepția feței (Michel et al., 1989; Sergent et al., 1992).,conectivitatea eficientă a fost testată de DCM-10, implementată în SPM8 toolbox (Wellcome Department of Imaging Neuroscience, Londra, Marea Britanie), care rulează sub Matlab R2008a (the MathWorks, Natick, MA, SUA). Modelele DCM sunt definite cu conexiuni endogene, reprezentând cuplarea între regiunile creierului (matricea A), conexiunile modulatoare (matricea B) și intrarea de conducere (matricea C). Aici, în matricea A, am definit conexiunile dintre regiunile selective ale feței (FFA și OFA) și LO., Imaginile fețelor și obiectelor au servit ca intrare de conducere (matricea C) și la această etapă de analiză nu am aplicat niciun efect modulator asupra conexiunilor.
estimarea modelului vizează maximizarea estimărilor negative de energie liberă ale modelelor (F) pentru un anumit set de date (Friston et al., 2003). Această metodă asigură că modelul fit utilizează parametrii într-un mod parcimonios (Ewbank et al., 2011). Modelele estimate au fost comparate, pe baza probelor modelului p (y / m), care este probabilitatea p de a obține date observate y date de un anumit model m (Friston et al., 2003; Stephan și colab., 2009)., În studiul de față aplicăm aproximarea negativă a energiei libere (variational free-energy) la dovezile jurnalului (MacKay, 2003; Friston et al., 2007). Selecția modelului Bayesian (BMS) a fost efectuată atât pe modelele de efecte aleatorii (RFX), cât și pe cele fixe (FFX) (Stephan et al., 2009). BMS RFX este mai rezistent la valori aberante decât FFX și nu presupune că același model ar explica funcția pentru fiecare participant (Stephan et al., 2009). Cu alte cuvinte, RFX este mai puțin sensibil la zgomot., În abordarea RFX rezultatul analizei este probabilitatea de depășire a spațiului modelului, care este măsura în care un model este mai probabil să explice datele măsurate decât alte modele. Cealaltă ieșire a analizei RFX este probabilitatea posterioară așteptată, care reflectă probabilitatea ca un model să genereze datele observate, permițând distribuții diferite pentru diferite modele. Ambele valori ale parametrilor sunt reduse prin lărgirea spațiului modelului (adică.,, prin creșterea numărului de modele), prin urmare, acestea se comportă într-o manieră relativă, iar modelele cu caracteristici comune și modele neplauzibile pot distorsiona rezultatul analizei. Prin urmare, pe lângă compararea directă a celor 28 de modele create, am împărțit spațiul modelului în familii, având modele de conectivitate similare, folosind metodele lui Penny et al. (2010).deoarece mai multe studii DCM anterioare indică legătura bidirecțională strânsă dintre FFA și OFA (Ishai, 2008; Gschwind et al.,, 2012) în analiza noastră, aceste două domenii au fost întotdeauna legate între ele și LO a fost conectat la ele în orice mod biologic plauzibil. Cele 28 de modele relevante au fost împărțite în trei familii de modele bazate pe diferențe structurale (Penny et al., 2010; Ewbank și colab., 2011). Familia 1 conține modele cu conexiuni liniare între cele trei zone, presupunând că informațiile curg de la LO la FFA prin OFA. Familia 2 conține modele cu o structură triunghiulară în care LO trimite intrări direct către FFA, iar OFA este, de asemenea, direct legată de FFA., Familia 3 conține modele, în care cele trei zone sunt interconectate, presupunând un flux circular de informații (Figura 2). Pentru a limita numărul de modele din această etapă a analizei, intrările au modulat exclusiv activitatea zonelor lor de intrare. Cele trei familii au fost comparate printr-un design aleatoriu BMS.
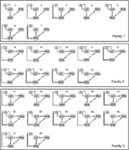
Figura 2. Cele 28 de modele analizate. Liniile negre marchează diviziunea dintre cele trei familii, având structuri diferite de matrice. Pentru detalii, consultați secțiunea ” Materiale și metode.,în al doilea rând, modelele din familia câștigătoare au fost elaborate în continuare prin crearea fiecărui model plauzibil cu conexiuni modulatoare, aplicând trei constrângeri. În primul rând, ambele fețe și obiecte modulează cel puțin o conexiune inter-areală. În al doilea rând, în cazul legăturilor bidirecționale, intrările modulatoare au un efect asupra ambelor direcții. În al treilea rând, dacă face dă o intrare directă în OFA, atunci presupunem că modulează întotdeauna conexiunea OFA-FFA (vezi Tabelul A2 din anexă). Aceste modele au fost introduse într-o a doua analiză BMS aleatorie din punct de vedere familial., În cele din urmă, membrii sub-familiei winner au fost înscriși într-un al treilea BMS pentru a găsi un singur model cu cea mai mare probabilitate de depășire.selecția modelului Bayesian a fost utilizată pentru a decide care Familie de modele explică cel mai bine datele măsurate. Ca rezultatele noastre arată cea de-a treia familie-a efectuat alte două, având o probabilitate de depășire de 0.995 comparativ cu primul familiei 0.00 și cea de-a doua familie este 0.004 (Figurile 3A,B)., Familia de modele câștigătoare (Familia 3; figura 2, partea de jos) conține 12 modele, având conexiuni între LO și FFA, OFA și FFA, și LO și OFA, dar care diferă atât în direcția conexiunilor, cât și în locul de intrare în rețea.

Figura 3. Rezultatele BMS RFX la nivelul familiilor. (A) probabilitățile așteptate ale comparației bazate pe familie sunt prezentate, cu probabilitățile comune de depășire (B).,ca o a doua etapă, toate modelele modulatoare posibile au fost proiectate pentru cele 12 modele din familia 3 (Vezi tabelul A2 din anexă; materiale și metode pentru detalii). Acest lucru a condus la 122 de modele, care au fost introduse în analiza aleatorie a familiei BMS, folosind 12 sub-familii. După cum se poate observa din figurile 4A,B, subfamilia model 4 a depășit celelalte subfamilii cu o probabilitate de depășire de 0,79. Ca o a treia etapă, modelele din sub-familia winner 4 au fost introduse într-un efect BMS aleatoriu. Figura 5 prezintă cele 18 variante testate ale modelului 20., Modelul cu cea mai mare probabilitate de depășire (p = 0,75) a fost modelul 4 (vezi Figura 6). Aceasta înseamnă că modelul winner conține conexiuni bidirecționale între toate zonele și intrările feței și obiectului, în mod surprinzător, ambele intră în LO. În plus, fețele au un efect modulator asupra legăturii dintre LO și FFA, în timp ce obiectele modulează legătura dintre Lo și OFA.

Figura 4. Rezultatele BMS RFX la nivelul sub-familiilor., (A) probabilitățile așteptate ale comparației bazate pe familie sunt prezentate, cu probabilitățile comune de depășire (B).

Figura 5. Modele de sub-familie 4. Toate modelele au același DCM.O structură (identică cu modelul 20 din familia 3), DCM.Structura B diferă de la model la model. Săgețile punctate au simbolizat efectul modulator al feței, în timp ce săgețile pătrate punctate arată modularea obiectului.
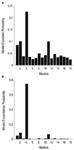
Figura 6., Rezultatele BMS în cadrul subfamiliei 4. (A) probabilitățile așteptate ale comparației bazate pe familie sunt prezentate, cu probabilitățile comune de depășire (B).pentru analiza estimărilor parametrilor modelului câștigător în grupul de subiecți a fost utilizată o aproximare aleatorie a efectului. Toate estimările maxime a posteriori (MAP) specifice subiectului au fost introduse într-un test t pentru mijloace unice și testate împotriva 0 (Stephan et al., 2010; Desseilles și colab., 2011). Rezultatele indexate cu un asterisc în Figura 7 diferă semnificativ de 0 (p < 0.,05).
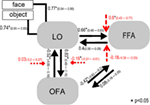
Figura 7. Structura modelului câștigător. Liniile Simple semnifică stimulii de intrare obiect și față în sistem (DCM.C). Săgețile negre arată conexiuni inter-regionale (DCM.A) în timp ce săgețile roșii reprezintă conexiunile modulatoare (DCM.B): modularea feței este semnificată cu săgeți punctate, în timp ce modularea obiectului este semnificată cu săgeți punctate cu cap pătrat. Mediile la nivel de grup ale estimărilor hărții și intervalele de încredere de 95% sunt ilustrate., Mediile au fost testate împotriva 0 și rezultatele semnificative sunt semnificate cu * if p < 0.05.,rezultatele majore ale prezentului studiu de conectivitate efectivă sugerează că (a) LO este legată direct de sistemul de procesare a feței OFA–FFA prin conexiuni bidirecționale cu ambele zone; (b) intrările non-față și față sunt amestecate la nivelul zonelor occipito-temporale și intră în sistem prin LO; (c) intrarea feței are un efect modulator asupra conexiunii LO și FFA, în timp ce intrările de intrarea modulează în mod semnificativ conexiunea Lo și ofa.rolul LO în percepția obiectelor este bine cunoscut din studiile anterioare (Malach et al.,, 1995; Grill-Spector et al., 1998a, b, 2000; Lerner și colab., 2001, 2008). Cu toate acestea, în ciuda studiilor anterioare, de obicei, a constatat o activitate crescută pentru obiecte complexe, precum și pentru fețele din LO, zona este de obicei asociată cu obiecte și o importanță relativ mai mică este atribuită rolului său în prelucrarea feței. În studiul de față, analiza eficientă a conectivității a poziționat LO în rețeaua centrală a percepției feței. Legătura directă dintre OFA și FFA a fost dovedită anterior atât din punct de vedere funcțional, cât și anatomic (Gschwind et al., 2012)., Cu toate acestea, deoarece nu există date actuale disponibile cu privire la rolul LO în acest sistem, l-am legat de celelalte două regiuni în mai multe moduri plauzibile.
primul BMS aleatoriu a arătat că în familia modelului câștigător LO este interconectat atât cu FFA, cât și cu OFA, iar conexiunile sunt bidirecționale. Prin urmare, subliniază faptul că LO poate avea o legătură structurală directă cu FFA., Cu modelarea tuturor efectelor modulatoare posibile, am constatat că a câștigat o sub-familie de modele, în care atât intrările de obiect, cât și cele de față intră în sistem prin intermediul LO, presupunând că LO joacă un rol general și important în regiunea de intrare. De la studiile anterioare de conectivitate funcțională, toate au început analiza rețelei de procesare a feței la nivelul IOG, corespunzând OFA (Fairhall și Ishai, 2007; Ishai, 2008; Cohen Kadosh et al., 2011; Dima și colab., 2011; Foley și colab., 2012) nu este surprinzător faptul că au trecut cu vederea rolul semnificativ al LO., Cu toate acestea, fețele sunt de fapt o categorie distinctă de obiecte vizuale, sugerând că neuronii sensibili la obiecte și forme ar trebui să fie activați, cel puțin într-o anumită măsură, și de fețe. Într-adevăr, studiile cu o singură celulă ale primatelor non-umane sugerează că cortexul inferior-temporal, homologul propus al LO uman în creierul macac (Denys et al., 2004; Sawamura și colab., 2006) are neuroni care răspund și la fețe (Perrett et al., 1982, 1985; Desimone și colab., 1984; Hasselmo și colab., 1989; Young și Yamane, 1992; Sugase și colab., 1999)., Legătura intimă a LO cu OFA și FFA, sugerată de prezentul studiu ar putea sublinia faptul că fețele și obiectele nu sunt prelucrate în întregime separat în calea vizuală ventrală, concluzie susținută și de datele imagistice funcționale recente (Rossion et al., 2012). Efectul modulator al intrării feței asupra conexiunii LO-FFA sugerează că LO trebuie să joace un rol în procesarea feței, cel mai probabil legat de prelucrarea structurală anterioară a fețelor, o sarcină pusă anterior în cea mai mare parte la OFA (Rotshtein et al., 2005; Fox și colab., 2009)., În cele din urmă, parametrii la nivel de grup ai modelului câștigător arată că legătura dintre lo, OFA și FFA este în principal ierarhică, iar conexiunile de feedback de la FFA către OFA și LO joacă un rol mai slab în sistem, cel puțin în timpul sarcinii de fixare aplicate.în concluzie, prin modelarea conectivității eficiente între zonele relevante pentru față și LO, sugerăm că LO joacă un rol semnificativ în prelucrarea fețelor.,
Declarație privind conflictul de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.această lucrare a fost susținută de Deutsche Forschungsgemeinschaft (KO 3918/1-1) și de Universitatea din Regensburg. Am dori să mulțumim Ingo Keck și Michael Schmitgen pentru discuții.Campbell, R. (2011). Speechreading și modelul Bruce-Young de recunoaștere a feței: constatări timpurii și evoluții recente. Br. J. Psychol. 102, 704–710.,
Pubmed Abstract | Pubmed Textul Complet | CrossRef Textul Complet
Fairhall, S. L., și Ishai, A. (2007). Conectivitate eficientă în cadrul rețelei corticale distribuite pentru percepția feței. Cereb. Cortex 17, 2400-2406.în 2000, Hoffman, E. A., and Haxby, J. V. (2000), a fost membru al Academiei de științe a Moldovei. Reprezentări distincte ale privirii și identității ochilor în sistemul neuronal uman distribuit pentru percepția feței. Nat. Neurosci. 3, 80–84.
Pubmed Abstract | Pubmed Textul Complet | CrossRef Textul Complet
Kourtzi, Z.,, și Kanwisher, N. (2001). Reprezentarea formei obiectului perceput de către complexul occipital lateral uman. Știință 293, 1506-1509.
Pubmed Abstract | Pubmed Textul Complet | CrossRef Textul Complet
MacKay, D. J. C. (2003). Teoria informației, inferență, și algoritmi de învățare. Cambridge: Cambridge University Press.
Nasanen, R. (1999). Lățimea de bandă de frecvență spațială utilizată în recunoașterea imaginilor faciale. Vision Res. 39, 3824-3833.
Pubmed Abstract | Pubmed Textul Complet | CrossRef Textul Complet
Rossion, B. Schiltz, C., și Crommelinck, M., (2003b). Occipitalul drept definit funcțional și „zonele feței” fusiforme discriminează romanul de fețele familiare vizual. Neuroimagine 19, 877-883.textul complet CrossRef
Sawamura, H., Orban, G. A. și Vogels, R. (2006). Selectivitatea adaptării neuronale nu corespunde selectivității răspunsului: un studiu cu o singură celulă al paradigmei de adaptare fMRI. Neuron 49, 307-318.
Pubmed Abstract | Pubmed Textul Complet | CrossRef Textul Complet
















Lasă un răspuns