Introducere
Cloroplastele reprezintă o caracteristică structurală a celulelor de plante și de sprijin de supraviețuire de plante prin metabolismul primar și funcții la nivel înalt (Kirk și Tilney-Bassett, 1978; Chefal, 1988; López-Juez și Pyke, 2005). În timpul creșterii vegetative a plantelor, celulele frunzelor conțin o populație foarte omogenă de cloroplaste în ceea ce privește dimensiunea și forma., Numărul de cloroplaste per celulă se realizează prin fisiune binară a pre-existente în organite și divizarea în două celule fiice in timpul diviziunii celulare (Birky, 1983; Possingham și Lawrence, 1983). Astfel, Reglarea numărului de cloroplaste într-un anumit tip de celulă este crucială pentru funcția celulară și moștenirea genetică a cloroplastelor.pentru a investiga natura determinării numărului de cloroplaste în celulele plantelor, celulele mezofile din frunze ale speciilor reprezentative au jucat un rol major ., Aceste celule sunt importante din punct de vedere fiziologic pentru fotosinteză și prezintă un grad ridicat de omogenitate structurală și funcțională. Mai devreme observarea sistematică analize de izolat țesuturi și de celule (Boasson și Laetsch, 1969; Possingham și Saurer, 1969; Boffey et al., 1979; Lamppa și colab.,, 1980; Thomas și Rose, 1983; Pyke și Lipitoare, 1991) a oferit multe informații utile pe cloroplastului numărul de determinare, inclusiv ideea că cloroplaste (plastide) nu sunt sintetizate de novo, dar reproduce prin diviziune și observația că frunza mesophyll cloroplastului număr este sensibil la diverse mediului și a plantelor-factori endogeni. Cu privire la acesta din urmă, în spanac, lumina are un impact pozitiv asupra cloroplastelor divizia timpul frunze de disc cultură comparativ cu dim sau condiții de întuneric (Possingham și Lawrence, 1983)., În primele frunze de grâu, volumul celular este corelat pozitiv cu proliferarea cloroplastelor (Ellis și Leech, 1985; Pyke și Leech, 1987). În Arabidopsis, fondul genetic afectează proliferarea cloroplastelor; numărul mediu de cloroplaste pe celulă în primele frunze este de 121 în Ecotipul Landsberg erecta (Ler) și 83 în ecotipul Wassilewskija (ws) (Pyke și Leech, 1994; Pyke și colab., 1994)., Frunze mesophyll celulele au, de asemenea, a contribuit la înțelegerea controlul genetic al cloroplastului divizare; de exemplu, screening-ul mutanți depreciate în cloroplast proliferarea și caracterizarea funcțiilor genetice implicate în această divizie au descoperit peste 20 de gene codifică cloroplastului divizia de componente de mașini sau cloroplastului factori de reglementare (Gao și Gao, 2011; Miyagishima et al., 2011; Basak și Møller, 2013; Osteryoung și Pyke, 2014; Li și colab., 2017).
prin contrast, studii privind replicarea cloroplastelor în celulele non-mezofile (de ex.,, celule de pavaj în epiderma frunzelor; Itoh și colab., 2018) sunt rare. Recent, s-a raportat că reglementarea diviziunii cloroplastelor diferă între țesuturile frunzelor (Fujiwara et al., 2018; Itoh și colab., 2018), deși mecanismul detaliat rămâne necunoscut. În plus, în timp ce analizele de suspensie-a cultivat DE-2 celule și frunze mesophyll protoplasts din tutun și trage meristem apical și frunze celulele primordiale în Arabidopsis (Nebenführ et al., 2000; Sheahan și colab.,, 2004; Seguí-Simarro și Staehelin, 2009) au oferit perspective majore, modul în care este reglementată partiționarea cloroplastului (plastid) în plante este încă neclar. Astfel, în ciuda eforturilor considerabile, rămân întrebări fundamentale în cercetarea cloroplastului, cum ar fi (i) cum este coordonat numărul cloroplastului pe celulă în țesuturile plantelor și (ii) cum este reglementată partiționarea cloroplastului la diviziunea celulară.,
Istoric al Cercetărilor privind Paza Celule Cloroplaste Număr
Stomatelor GCs în epiderma trage, în general, conțin cloroplaste și control schimbul de gaze între frunze mesophyll și atmosferă (Sachs, 1875; Taiz et al., 2015; a se vedea figura 1A). Prima investigație a numărului de cloroplast GC în frunze a fost efectuată în urmă cu peste un secol în plantele Drosera cultivate natural (Macfarlane, 1898)., Acest studiu a demonstrat că, ca și alte caracteristici structurale ale plantelor și celulelor, numărul GC cloroplast pe celulă într-un hibrid presupus derivat dintr-o încrucișare între Drosera filiformis și Drosera intermedia a fost intermediar între cele două specii, ceea ce implică faptul că numărul GC cloroplast ar putea fi utilizat pentru a determina structura genetică a unei plante. Observații importante au fost raportate ulterior cu privire la diferențele în numărul de cloroplast GC între speciile de plante (Sakisaka, 1929) și numărul relativ stabil de cloroplast în GCs în epiderma frunzelor de dud (Morus spp.,; Hamada și Baba, 1930) și în frunze mature de mai multe specii de Brassica (Iura, 1934). În plus, analiza de autopolyploid sfeclă de zahăr (Beta vulgaris) plante dezvăluit că GC numărul cloroplastelor din frunze este corelat pozitiv cu nuclear nivelul de ploidie al plantelor (Mochizuki și Sueoka, 1955). Mai multe analize aprofundate și cuprinzătoare au fost apoi efectuate folosind diferite probe de plante pentru a investiga relația dintre numărul de cloroplast și dimensiunea stomatală cu nivelul ploidy (de exemplu, Frandsen, 1968)., În aceste analize, cloroplast de numărare la stomatelor (GC pereche) nivel a fost frecvent adoptate, care exclude efectul de părtinitoare cloroplast de distribuție între asociat GCs (de exemplu, Mochizuki și Sueoka, 1955; Frandsen, 1968), dezvăluind că media GC numărul cloroplastelor din frunze sau cotiledoane în aproximativ 80 de specii, variante, sau hibrizi au variat de la 2,8 la 40.0 în diploids (2×) și 5.0 la 73.5 în tetraploids (4×). În plus, evenimentele de duplicare a întregului genom în plante (de exemplu, 1× la 2×, 2× la 4× etc.) a provocat o aproximativ 1.,Creșterea de 7 ori a numărului de cloroplast GC cu fidelitate ridicată (revizuită în Butterfass, 1973). Aceste rezultate încurajat investigații în ploidie nivel în diferite țesuturi și plante obținute prin culturi de țesuturi, de trecere, naturale sau de cultivare, în combinație cu substanțe chimice (de exemplu, colchicina) sau tratamente cu radiații (de exemplu, Jacobs și Yoder, 1989; Singsit și Veilleux, 1991; Qin și Rotino, 1995)., În timp ce GC cloroplastului număr a fost studiat în stomatelor biologie (Lawson, 2009) și citologie pentru a înțelege această multiplicare (Butterfass, 1979; a se vedea mai jos), are, în mare măsură, a servit ca un fiabile și convenabil marker pentru detectarea de hibrizi, dintre specii și variante și pentru estimarea numărului de niveluri de țesuturi vegetale țintă.
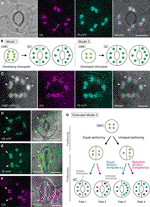
Figura 1 modele Simplificate și microscopie de probe pentru controlul cloroplastului numărul stomatelor garda de celule (GCs)., (A) o stomă tipică (pereche GC) în epiderma abaxială a lamei de frunze Arabidopsis care exprimă o fuziune orientată spre stomă a peptidei de tranzit (TP) cu proteina fluorescentă ciană (PCP; TP-PCP). (B) două modele de determinare a numărului de cloroplast în GCs, care implică fie partiționarea cloroplastelor (modelul 1), fie proliferarea și partiționarea cloroplastelor (modelul 2) în timpul dezvoltării GC din celulele mamă de protecție (GMC). (C) o pereche de GC în epiderma adaxială a Pețiolului frunzelor de Arabidopsis care exprimă TP-CFP și FtsZ1 fuzionată cu proteina fluorescentă verde (GFP; FtsZ1-GFP)., (D–f) perechi GC în epidermă abaxială a lamei de frunze Arabidopsis cu (D, E) sau fără (F) expresia TP fuzionată cu proteina fluorescentă galbenă (YFP; TP-YFP). (F) Autofluorescența clorofilei (Chl) a fost utilizată ca marker de cloroplast., (G) a Extins modelul 2, care reprezintă implicarea egale și inegale cloroplastului partiționare următoarele GMC divizie și, ulterior, divizia de GC cloroplaste cu egale (linia albastră) sau selectivă (linia roșie) divizia de competență, care ar conduce la patru tipuri de cloroplaste numărul de determinare (Soarta 1-4) în timpul târziu stomatelor dezvoltarea de Arabidopsis frunze. (A, C–F) microscopia cu Epifluorescență a fost efectuată cu un microscop inversat Olympus IX71 folosind materiale vegetale așa cum s-a descris anterior (Fujiwara et al., 2017, Fujiwara și colab., 2018)., Semnalele fluorescente ale clorofilei, CFP, GFP și YFP sunt pseudo-colorate în magenta, cyan, Verde (numai în imagine fuzionată) și, respectiv, verde. Indicațiile din panouri sunt următoarele: capul săgeții, inelul FtsZ1; săgeată, cloroplast GC mărit; linie punctată, formă celulară. Bara de scară = 10 µm.
utilitatea celulelor de protecție a frunzelor pentru analiza controlului numărului de cloroplast
celulele mezofile din frunze au fost folosite de mult timp ca model primar pentru analiza numărului de cloroplast., În timp ce acestea au avantaje pentru studiul efectelor condițiilor de mediu asupra cloroplastelor diviziune (de exemplu, lumina-regulator de semnalizare; Boasson și Laetsch, 1969; Possingham și Lawrence, 1983; Okazaki et al., 2009; Chiang și colab., 2012), acestea sunt limitate în anumite privințe. În primul rând, celulele mezofile din frunze variază ca mărime și formă și sunt distribuite adânc în interiorul frunzei, ceea ce face dificilă manipularea țesuturilor intacte., În al doilea rând, susceptibilitatea proliferării mezofilei cloroplastelor la stresul de mediu și condițiile de creștere a plantelor poate împiedica comparații fiabile între studii. Numărul de mesofil cloroplast pe celulă în Ecotipul Arabidopsis Columbia (Col) a fost raportat diferit ca 76 (Kinsman și Pyke, 1998), 80-100 (Stokes et al., 2000), 70 (Tirlapur și König, 2001), 41 (Yoder și colab., 2007), 40-60 (Okazaki et al., 2009)și 30-40 (Kawade et al., 2013)., În plus, este aproape imposibil să se evalueze contribuția partiționării cloroplastului la numărul final de cloroplast pe celulă mezofilă în timpul dezvoltării frunzelor, deși se consideră că acest lucru este determinat de echilibrul dintre rata diviziunii celulare și rata diviziunii cloroplastului. Pentru a descoperi mecanismul de control al numărului de cloroplast în celulele frunzelor vegetative, este necesar un sistem model care depășește problemele de mai sus.GCs Stomatal (vezi figura 1a) prezintă caracteristicile unui sistem model pentru înțelegerea mecanismului de control al numărului de cloroplast., GCs sunt foarte uniforme în dimensiune și formă într-un țesut, iar distribuția lor împrăștiată, dar densă, în stratul exterior al lăstarilor facilitează detectarea lor prin microscopie de lumină și fluorescență. GCs sunt, de asemenea, derivat din protodermal celulele trage meristem apical sau din embrionare celule epidermice, și lor secvență de dezvoltare prin meristemoids (a stomatelor precursor cu meristematice activitate) și garda mama celule (GMCs; un precursor al GC perechi) este stabilită în detaliu (Zhao și Sac, 1999; Nadeau și Sac, 2002; Kalve et al., 2014)., Dezvoltarea stomatologică tardivă implică o singură rundă de divizare simetrică a GMC, care permite evaluarea distribuției și partiționării cloroplastelor înainte și după citokineză. Din perspectiva experimentelor practice, GCS-urile de frunze sunt potrivite pentru microscopie. S-a demonstrat anterior că numărul de cloroplast pe celulă din GCs de frunze de Sinapis alba a fost mai puțin afectat de condiții de lumină diferite decât cel din celulele mezofile de frunze (Wild și Wolf, 1980)., În plus, diferența dintre numărul de cloroplast GC în pețiolele frunzelor este relativ minoră între cele trei ecotipuri Arabidopsis Col, ler și Ws (Fujiwara et al., 2018). În plus, endoreduplicarea, care are impact asupra dezvoltării mezofilei frunzelor, a pavajului și a celulelor trichome, nu a fost detectată în GCS de frunze Arabidopsis (Melaragno et al., 1993), care ar asigura interpretările datelor numărului de cloroplast la nivelul 2c al celulelor., Împreună, aceste rapoarte sugerează că GCS leaf sunt potențial un model excelent pentru analiza sistematică a dinamicii numărului de cloroplast într-o anumită linie celulară.
Arabidopsis Leaf Guard Cells ca Model pentru studierea controlului numărului de cloroplast
în istoria cercetării GC chloroplast, numărarea cloroplastelor la nivelul stomatal (GC pair) a servit un rol la fel de important în determinarea numărului de cloroplast ca numărarea la nivelul GC individual. Ambele metode produc același număr mediu de cloroplast (Butterfass, 1973)., Atunci când variația distribuției cloroplastelor în GCs pereche și mecanismul său de bază este un subiect de focalizare, este esențială informația detaliată a cloroplastelor la nivelul celulei individuale, adică dimensiunea, forma și localizarea intracelulară a acestora. Cloroplast (plastid) proliferarea timpul GMC–GC diferențiere a fost anterior cercetat în mai multe specii de plante (Butterfass, 1973, Butterfass 1979)., Aceste studii au propus două modele pentru determinarea numărului terminal de cloroplast în GCs la diferite specii de plante (figura 1b): unul (model 1; sfeclă de zahăr) implică doar partiționarea cloroplastului la diviziunea GMC, iar celălalt implică nu numai partiționarea cloroplastului, ci și proliferarea cloroplastului în timpul dezvoltării GC.în epoca geneticii moleculare, genomicii, imagisticii celulare și a altor analize interdisciplinare, există multe posibilități pentru caracterizarea ulterioară a mecanismului de partiționare a cloroplastului., Arabidopsis leaf GCS poate fi unul dintre cele mai bune sisteme de model în acest scop. Mai multe studii au examinat numărul de cloroplast GC în frunzele sau cotiledoanele Arabidopsis (Hoffmann, 1968; Pyke și Leech, 1994; Pyke și colab., 1994; Robertson și colab., 1995; Keech și colab., 2007; Chen și colab., 2009; Yu și colab., 2009; Higaki și colab., 2012; Fujiwara și colab., 2018). Aceste GCs prezintă un număr modest de cloroplaste, variind de la 3,5 la 5,5 în medie. Până în prezent, niciun studiu nu a examinat modificările numărului de cloroplast (plastid) în timpul dezvoltării stomatale., Cu toate acestea, dovezile microscopice din analizele dezvoltării stomatale (de exemplu, Zhao și Sack, 1999; Hachez et al., 2011) și observații preliminare indică faptul că GMCs poate conține un număr mai mic de a dezvolta cloroplaste decât GCs și că această proliferare pot apărea în timpul GC diferențiere. Pentru a testa acest lucru, formarea cloroplastelor divizia de mașini în GCs a fost monitorizată cu sonda FtsZ1 fuzionat la proteina fluorescentă verde (FtsZ1-GFP) (Fujiwara et al., 2008)., Un transgenice linie, simultan exprima un tranzit peptide (TP)-punte cu PCP și FtsZ1-GFP pentru a vizualiza stroma și FtsZ1 inel, respectiv, a fost examinat de epifluorescenta microscopie (Fujiwara et al., 2017). Extinderea pețiolelor frunzelor (a cincea frunze de răsaduri vechi de 4 săptămâni) au fost folosite. Ca rezultat, s-au detectat GCs cu cloroplaste constrictive simetric (figura 1C). Aceste cloroplaste format FtsZ1 inel, un cloroplast inel cu diviziune de pe stromale suprafața plicului interior membrana din frunze mesophyll și trotuar celule (Vitha et al., 2001; Fujiwara și colab.,, 2008), la locul constricției ecuatoriale. În concordanță cu modelul stomatal în dezvoltarea frunzelor Arabidopsis (Donnelly et al., 1999; Andriankaja și colab., 2012), cloroplastele de divizare au fost detectate atât la etapa târzie, cât și la cea timpurie a expansiunii frunzelor. Astfel, modelul 2 este cel mai probabil cel mai potrivit pentru Arabidopsis leaf GCS.
relația dintre proliferarea cloroplastelor și expansiunea în celulele de protecție a frunzelor Arabidopsis
În plus, a fost observat un fenotip neașteptat de morfogeneză a cloroplastelor GC la GCs mature (figurile 1d, E)., Când cojile epidermice ale frunzelor complet extinse (lamele a treia-a patra a frunzelor de răsaduri vechi de 4 săptămâni) dintr-o linie de proteine fluorescente galbene topite TP (YFP) au fost caracterizate microscopic (linia FL6-5; Fujiwara et al., 2018), unele stomate au prezentat modele inegale de distribuție a cloroplastelor în perechi GC, în timp ce majoritatea stomatelor frunzelor au prezentat modele de distribuție a cloroplastelor egale sau similare (Robertson et al., 1995; Fujiwara și colab., 2018)., În cadrul perechii GC a unei stome, Mărimea cloroplastelor din GC care conține un număr mai mic de cloroplaste a fost mai mare decât în celelalte GCs din perechea care conține un număr mai mare de cloroplaste (figurile 1d, E). În acest fel, GCs menține probabil volumul total de cloroplast pe celulă la un nivel constant în timpul creșterii celulare. Cloroplastele lărgite reprezentau fenotipul terminal și nu se mai puteau împărți în frunze expandate. Aceste rezultate au fost confirmate în mai multe experimente independente, indiferent de expresia unei proteine fluorescente topite cu TP pentru etichetarea stromei (figura 1F).,acest fenotip de cloroplast GC este interpretat ca un mecanism de compensare a expansiunii cloroplastului, care a fost bine documentat în celulele mezofile din frunze defecte în controlul diviziunii cloroplastului (Pyke și Leech, 1994; Pyke și colab., 1994). Până în prezent, un singur studiu (Ellis și Lipitoare, 1985) a raportat o corelație negativă între cloroplast numărul și mărimea cloroplastelor din frunze mesophyll celule de grâu, întrucât numeroase studii au raportat o corelație pozitivă între volumul celulelor și numărul cloroplastelor din frunze normale mesophyll celule (Leech și Pyke, 1988; Pyke, 1997)., Întrucât dezechilibrele în numărul de cloroplast GC apar la frecvență joasă (Fujiwara et al., 2018), eterogenitatea cloroplastului în perechile GC indică faptul că partiționarea inegală a cloroplastului ar putea declanșa o creștere diferențială a cloroplastului între celulele frunzelor de tip sălbatic din Arabidopsis, în ciuda diviziunii celulare simetrice.efectul de compensare a cloroplastului în GCs poate fi mai puțin strict decât în celulele mezofile din frunze., GCs-ar putea fi capabil să reziste la deficitul sau pierderea completă a total cloroplastului volum per celulă în insuficiență cloroplastului divizia mutanți, cum ar fi în Arabidopsis arc6 și atminE1 și roșii suffulta, întrucât multe mutant GCs-a arătat redus numărul cloroplastelor și extinsă această dimensiune în mod similar cu frunze mesophyll celule (Robertson et al., 1995; Forth și Pyke, 2006; Chen și colab., 2009; Fujiwara și colab., 2018)., Într-un târziu cloroplastului divizia mutant, arc5, reducerea GC cloroplastului număr nu a fost asociată cu o creștere semnificativă în această dimensiune, spre deosebire de frunze mesophyll celule (Pyke și Lipitoare, 1994). Un grad mai mic de expansiune a cloroplastului în GCs decât în celulele mezofile (Pyke și Leech, 1994; Barton și colab., 2016) și variația expansiunii cloroplastului în rândul GCs, ar putea sta la baza unei game atât de largi admise de volum total de cloroplast pe GC., În plus, momentul diviziunii cloroplastului în timpul diferențierii GMC–GC ar putea afecta în mod semnificativ fenotipul terminal al cloroplastului GC. Deși este necesară o caracterizare detaliată suplimentară pentru a aborda această problemă, pare plauzibil faptul că GCS de frunze Arabidopsis reprezintă un sistem de investigare a aspectelor neexploatate ale controlului numărului de cloroplast în celulele plantelor.,
un Model de lucru pentru determinarea numărului de cloroplast în celulele de protecție a frunzelor Arabidopsis
pe baza celor de mai sus, propunem un model de lucru (un model extins 2) pentru analiza numărului de cloroplast în GCs (figura 1g). Numărul final de cloroplast pe GC este determinat prin partiționarea cloroplastului la diviziunea GMC și proliferarea cloroplastului în GCs. În timpul diviziunii GMC, cloroplastele pot suferi o partiționare egală sau inegală. În timpul proliferării cloroplastelor, cloroplastele GC vor prolifera fie cu o competență de diviziune egală (linia albastră), fie selectivă (linia magenta)., De exemplu, dacă cloroplastele împărțite în mod egal posedă o competență de divizare echivalentă, numerele egale de cloroplast vor apărea în perechea GC (soarta 1). Dacă cloroplastele partiționate inegal posedă o competență de divizare echivalentă, cloroplastele vor crește cu aceeași rată în cadrul perechii GC (Fate 3). Dacă diviziunea selectivă a cloroplastului are loc în GCs, soldul numărului de cloroplast din perechea GC se va schimba după diviziunea GMC (destinele 2 și 4)., În prezent, este dificil să se găsească sprijin pentru „diviziunea selectivă a cloroplastelor”, dar dacă destinele 1 și 4 predomină de fapt în GCs, atunci acestea ar putea avea un mecanism care controlează volumul total de cloroplast pe celulă, ca în celulele mezofile din frunze. Modelul ridică două probleme: (i) cloroplastele GC sunt împărțite corespunzător în celule fiice și cum se partiționează? Și (ii) competența de divizare a cloroplastelor GC este reglementată în mod coordonat?,
în ceea ce Privește problema (i), dacă această moștenire are loc prin repartizare aleatorie de mai multe cloroplaste în citoplasmă sau prin pozitive cloroplastului partiționare mecanism(e) a fost o veche preocupare (Butterfass, 1969; Birky, 1983; Hennis și Birky, 1984; Nebenführ, 2007; Sheahan et al., 2016). Surprinzător, în Arabidopsis arc6, frunze sau cotiledon GCs au zero la trei cloroplaste, și în cloroplast-deficit de GCs, non-plastide fotosintetic există încă în veziculoasă la forme alungite (Robertson et al., 1995; Chen și colab., 2009; Fujiwara și colab., 2018)., Nu GCs lipsite de plastide per se a fost găsit în arc6, și nici o explicație pentru acest lucru a fost viitoare, în ciuda perturbare a cloroplastului divizia de aparate (Vitha et al., 2003). În consecință, va fi important să se examineze replicarea și morfologia cloroplastelor arc6 în studiile de linie stomatală. Arabidopsis mutant de cercetare poate da, de asemenea, un alt indiciu pentru această problemă., Observația că 18% din cotiledon GCs în frunze mototolită (crl) mutant nu conțin plastidic structuri în citoplasmă, în timp ce 100% din frunze de mesophyll celulele conțin una la patru extinsă cloroplaste (Asano et al., 2004; Chen și colab., 2009), este de mare importanță. CRL este o proteină cu înveliș exterior de cloroplast cu o funcție necunoscută. Înțelegerea funcției proteinei CRL poate oferi informații despre mecanismul(mecanismele) de partiționare cu cloroplast., Analiza de cloroplast proliferarea și partiționare în frunze mesophyll celule în Arabidopsis arc mutanți și alte linii transgenice a promovat cercetarea în proliferarea și de partiționare de non-mesophyll plastide. De asemenea, rezultatele obținute în GCs pot fi transferabile către alte sisteme celulare.
observație finală
modelul GC deschide multe perspective pentru dezvoltarea biologiei cloroplastelor. De exemplu, în timp ce sistemele citoscheletice sunt cunoscute pentru a regla morfologia cloroplastului, mișcarea și partiționarea (Sheahan et al.,, 2016; Wada, 2016; Erickson și Schattat, 2018), rolul fiecărei gene de reglementare în proliferarea și partiționarea cloroplastelor în plante a primit puțină atenție. Pe de altă parte, odată ce devine posibil să afecteze GC numărul cloroplastelor sau morfologie prin diferite strategii experimentale, perspective noi în molecular de control al cloroplastului morfogeneza în stomatelor lineage celulele pot fi furnizate. În plus, în combinație cu analizele cantitative ale numărului de cloroplast în timpul dezvoltării stomatale, modelarea matematică poate oferi o nouă cale pentru aceste investigații., Această lucrare prezintă cunoștințele actuale despre modul în care este controlat numărul GC chloroplast și evidențiază utilitatea potențială a GCS de frunze Arabidopsis pentru înțelegerea proliferării și partiționării cloroplastului.
Declarația de disponibilitate a datelor
toate seturile de date pentru acest studiu sunt incluse în articol/ materialul suplimentar.
contribuții ale autorului
MF a conceput studiul și a scris manuscrisul. MF și AS au efectuat experimentele. AS și RI au efectuat analizele. Toți autorii au citit și aprobat manuscrisul final.,
de Finanțare
Această lucrare a fost susținută de către Ministerul Educației, Culturii, Științei și Tehnologiei din Japonia sub KAKENHI (grant nr. 19K05831 la MF și 18K06314 a RI).
Conflict de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
mulțumiri
autorii mulțumesc Dr. Nobuyuki Kanzawa (Universitatea Sophia) pentru sfaturi cu privire la activitatea.
Birky, C. W., Jr. (1983)., Partiționarea organite citoplasmatice la diviziunea celulară. Int. Rev. Cytol. 15, 49-86. Google Scholar
butter putinei, T. (1969). Distribuția plastidă în mitoză a celulelor mamă de închidere a haploid swedenklee (Trifolium hybridum L.). Planta 84, 230-234. doi: 10.1007/BF00388108
PubMed Abstract | CrossRef Textul Complet | Google Scholar
unt putinei, T. (1973). Controlul diviziunii plastide prin intermediul cantității de ADN nuclear. Protoplasma 76, 167-195. doi: 10.,1007/BF01280696
CrossRef Full Text | Google Scholar
Qin, X., Rotino, G. L. (1995). Chloroplast number in guard cells as ploidy indicator of in vitro-grown androgenic pepper plantlets. Plant Cell Tissue Organ Cult. 41, 145–149. doi: 10.1007/BF00051583
CrossRef Full Text | Google Scholar
Sachs, J. (1875). Textbook of Botany. Oxford: Clarendon Press.
Google Scholar
Thomas, M. R., Rose, R. J. (1983)., Numărul Plastid și modificările structurale plastid asociate cu cultura protoplastului mezofil de tutun și regenerarea plantelor. Planta 158, 329-338. doi: 10.1007/BF00397335
PubMed Abstract | CrossRef Textul Complet | Google Scholar
Tirlapur, U. K., König, K. (2001). Lasere femtosecunde cu infraroșu apropiat ca un instrument nou pentru imagistica non-invazivă în timp real, de înaltă rezoluție, a diviziunii cloroplastului în celulele vii de teacă de pachete de Arabidopsis. Planta 214, 1-10. doi: 10.,1007/s004250100597
PubMed Abstract | CrossRef Textul Complet | Google Scholar
Sălbatice, A., Lup, G. (1980). Efectul diferitelor intensități ale luminii asupra frecvenței și mărimii stomatelor, mărimii celulelor, numărului, mărimii și conținutului de clorofilă a cloroplastelor în mezofilă și celulele de gardă în timpul ontogeniei frunzelor primare de Sinapis alba. Z. Pflanzenfiziol. Bd. 97, 325–342. doi: 10.1016/S0044-328X(80)80006-7
CrossRef Textul Complet | Google Scholar
















Lasă un răspuns