Inleiding
de neurale verwerking van gezichten is een veel onderzocht onderwerp van de cognitieve wetenschap. Op basis van functionele beeldvormingsstudies, single-cell opnames en neuropsychologisch onderzoek is gesuggereerd dat face-processing wordt uitgevoerd door een gedistribueerd netwerk, waarbij verschillende corticale gebieden van de zoogdierhersenen betrokken zijn (Haxby et al., 2000; Marotta et al., 2001; Rossion et al., 2003a; Avidan et al., 2005; Sorger et al., 2007)., Hoewel de omvang van dit face-processing netwerk momenteel onder intensief debat (Ishai, 2008; Wiggett and Downing, 2008) de meeste onderzoekers het erover eens dat er tal van corticale gebieden geactiveerd door face stimuli. The most invloedrijk model of face perception, based on the original model of Bruce and Young, 1986; Young and Bruce, 2011) stelt een onderscheid voor tussen de representatie van invariante en variante aspecten van face perception op een relatief onafhankelijke manier, gescheiden in een “core” en een” extended ” deel (Haxby et al., 2000)., De belangrijkste gebieden van het” kernnetwerk ” zijn gebieden van het occipitale en de laterale spoelvormige gyri (FG). De gebieden van deze twee anatomische regio ‘ s lijken te zijn gespecialiseerd voor verschillende taken: terwijl het occipitale gezicht gebied (OFA), gelegen op de inferieure occipitale gyrus (IOG) lijkt te zijn betrokken bij de structurele verwerking van gezichten, de spoelvormige gezicht gebied (FFA) processen geconfronteerd op een hoger niveau, bij voorbeeld bij de verwerking van identiteit (Sergent et al., 1992; George et al., 1999; Ishai et al., 1999; Hoffman and Haxby, 2000; Rossion et al., 2003a, b; Rotshtein et al.,, 2005). Daarnaast zijn de veranderlijke aspecten van gezichten (zoals gezichtsuitdrukkingen, richting van Oog-Blik, expressie, lip bewegingen (Perrett et al., 1985, 1990), of liplezen (Campbell, 2011) lijken te worden verwerkt in de superieure temporale sulcus (STS; Puce et al., 1998; Hoffman and Haxby, 2000; Winston et al., 2004). De drie bovengenoemde gebieden (FFA, OFA, STS) vormen de zogenaamde “kern” van het perceptuele systeem van gezichtsverwerking (Haxby et al., 2000; Ishai et al., 2005)., Terwijl basisinformatie over gezichten door dit kernsysteem wordt verwerkt, voegt complexe informatie over de stemming van de anderen, het niveau van interesse, aantrekkelijkheid of richting van aandacht ook informatie toe aan gezichtsperceptie en wordt verwerkt door een extra, zogenaamd “uitgebreid” systeem (Haxby et al., 2000). Dit systeem bevat hersengebieden met een grote verscheidenheid aan cognitieve functies met betrekking tot de verwerking van veranderlijke gezichtsaspecten (Haxby et al., 2000; Ishai et al., 2005) en omvatten gebieden zoals de amygdala, insula, de inferieure frontale gyrus evenals de orbitofrontale cortex (Haxby et al.,, 2000, 2002; Fairhall and Ishai, 2007; Ishai, 2008).
interacties van de bovengenoemde gebieden worden in dit onderzoek gemodelleerd met behulp van methoden die de effectieve connectiviteit tussen corticale gebieden berekenen. Dynamische causale modellering (DCM) is een veel gebruikte methode om effectieve connectiviteit tussen hersengebieden te verkennen. Het is een generieke benadering voor het modelleren van de wederzijdse invloed van verschillende hersengebieden op elkaar, gebaseerd op fMRI-activiteit (Friston et al., 2003; Stephan et al., 2010) en om het verbindingspatroon van corticale gebieden in te schatten., DCMs zijn generatieve modellen van neurale reacties, die “a posteriori” schattingen van synaptische verbindingen tussen neuronale populaties (Friston et al., 2003, 2007). Het bestaan van het gedistribueerde netwerk voor face-processing werd voor het eerst bevestigd door functionele connectiviteitsanalyse door Ishai (2008), die beweerde dat het centrale knooppunt van face-processing de laterale FG is, verbonden met lagere-orde gebieden van de IOG evenals met STS, amygdala, en frontale gebieden. De eerste poging om het face-processing netwerk te onthullen in het geval van realistische dynamische gezichtsuitdrukkingen werd gedaan door Foley et al. (2012)., Zij bevestigden de rol van IOG, STS, en FG gebieden en vonden dat de verbindingssterkte tussen leden van het kernnetwerk (OFA en STS) en van het uitgebreide systeem (amygdala) worden verhoogd voor het verwerken van beà nvloede gebaren. Momenteel hebben verschillende studies ons begrip van het face-processing netwerk uitgewerkt en een directe link tussen amygdala en FFA en de rol van deze verbinding in de perceptie van angstige gezichten (Morris et al., 1996; Marco et al., 2006; Herrington et al., 2011)., Ten slotte werd het effect van hogere cognitieve functies op gezichtswaarneming ook gemodelleerd door de verbindingen van de orbitofrontale cortex met het kernnetwerk te testen (Li et al., 2010). Het werd gevonden (Li et al., 2010) dat de orbitofrontale cortex een effect heeft op de OFA, wat de informatieverwerking van de FFA verder moduleert.
hoewel eerdere effectieve connectiviteitsstudies de details van het gezichtsverwerkings netwerk met betrekking tot verschillende aspecten van gezichtsperceptie aan het licht brachten, werd het simpele feit genegeerd dat gezichten ook als visuele objecten kunnen worden beschouwd., We weten uit een groot aantal experimenten dat visuele objecten worden verwerkt door een gedistribueerd corticaal netwerk, met inbegrip van vroege visuele gebieden, occipito-temporale en ventrale–temporale cortices, grotendeels overlappen met het face-processing netwerk (Haxby et al., 1999, 2000, 2002; Kourtzi en Kanwisher, 2001; Ishai et al., 2005; Gobbini and Haxby, 2006, 2007; Haxby, 2006; Ishai, 2008). Een van de belangrijkste gebieden van visuele objectverwerking is de laterale occipitale cortex (LOC), die kan worden onderverdeeld in twee delen: de anterior–ventral (PF/LOa) en de caudal–dorsal deel (LO; Grill-Spector et al.,, 1999; Halgren et al., 1999). De LOC werd voor het eerst beschreven door Malach et al. (1995), die verhoogde activiteit gemeten voor objecten, waaronder beroemde gezichten ook, in vergelijking met roerei objecten (Malach et al., 1995; Grill-Spector et al., 1998a). Sindsdien wordt het laterale occipitale (LO) voornamelijk beschouwd als een object-selectief gebied, dat niettemin steevast ook verhoogde activering voor gezichten heeft (Malach et al., 1995; Puce et al., 1995; Lerner et al., 2001), vooral voor omgekeerde (Aguirre et al., 1999; Haxby et al., 1999; Epstein et al.,, 2005; Yovel and Kanwisher, 2005).het is dan ook nogal verwonderlijk dat, hoewel verscheidene studies de effectieve connectiviteit van frontverwerkingsgebieden hebben behandeld, geen van deze studies de rol van de LO in het netwerk in aanmerking heeft genomen. In een eerdere fMRI-studie vonden we dat LO een cruciale rol speelt in de zintuiglijke competitie voor gezichtsprikkels (Nagy et al., 2011)., De activiteit van LO werd verminderd door de presentatie van gelijktijdig gepresenteerde gelijktijdige stimuli en deze responsreductie, die zintuiglijke competitie tussen stimuli weerspiegelt, was groter wanneer de omringende stimulus een gezicht was in vergelijking met een Fourier-fase gerandomiseerd ruisbeeld. Dit resultaat ondersteunde ook het idee dat LO een specifieke rol kan spelen in gezichtswaarneming. Daarom hebben we hier expliciet onderzocht, met behulp van methoden van effectieve connectiviteit, hoe LO is gekoppeld aan FFA en OFA, leden van het voorgestelde kernnetwerk van gezichtsperceptie (Haxby et al., 1999, 2000, 2002; Ishai et al., 2005).,
materialen en methoden
proefpersonen
vijfentwintig gezonde deelnemers namen deel aan het experiment (11 vrouwen, mediaan: 23 jaar, min.: 19 jaar, max.: 35 jaar). Ze hadden allemaal een normaal of gecorrigeerd gezichtsvermogen( zelf gemeld), geen van hen had neurologische of psychische aandoeningen. Onderwerpen die hun schriftelijke geïnformeerde toestemming in overeenstemming met de protocollen goedgekeurd door de Ethische Commissie van de Universiteit van Regensburg.,
Stimuli
proefpersonen werden centraal gepresenteerd door grijsschaalvlakken, niet-zintuiglijke objecten en de Fourier gerandomiseerde versies van deze stimuli, gecreëerd door een algoritme (Nasanen, 1999) dat het fasespectrum vervangt door willekeurige waarden (variërend van 0° tot 360°), waardoor het amplitudespectrum van het beeld intact blijft, terwijl alle vorminformatie wordt verwijderd. Gezichten waren full-front digitale beelden van 20 jonge mannen en 20 jonge vrouwen. Ze zaten achter een rond masker (3,5 ° diameter) dat de buitenste contouren van de gezichten elimineerde (zie een voorbeeld in Figuur 1)., Objecten waren niet-sense, gerenderde objecten (n = 40) met dezelfde gemiddelde grootte als het gezichtsmasker. De luminantie en het contrast (d.w.z. de standaardafwijking van de luminantieverdeling) van de stimuli werden gelijkgesteld door de luminantie histogrammen (gemiddelde Luminantie: 18 cd/m2) met Photoshop aan te passen. Stimuli werden via een LCD-videoprojector (JVC, DLA-G20, Yokohama, Japan, 72 Hz, 800 × 600 resolutie) terug geprojecteerd op een doorschijnend circulair scherm (app. 30 ° diameter), geplaatst in de scanner boring op 63 cm van de waarnemer., Stimulus presentatie werd gecontroleerd via E-prime software (Psychological Software Tools, Pittsburgh, PA, USA). Gezichten, objecten en Fourier ruisbeelden werden gepresenteerd in volgende blokken van 20 s, interleaved met 20 s van blanco perioden (uniforme grijze achtergrond met een luminantie van 18 cd/m2). Stimuli werden gepresenteerd voor 300 ms en werden gevolgd door een isi van 200 ms (2 Hz) in een willekeurige volgorde. Elk blok werd vijf keer herhaald. De deelnemers werd gevraagd zich continu te concentreren op een centraal gepresenteerd fixatieteken., Deze functionele localizer runs maakten deel uit van twee andere experimenten van gezicht perceptie, elders gepubliceerd (Nagy et al., 2009, 2011).

figuur 1. Proefprikkels van het experiment. Alle beelden waren grijsschaal, hetzelfde in grootte, luminantie en contrast. Linkerpaneel toont een gezicht, geslacht specifieke kenmerken (zoals haar, sieraden etc.) was verborgen achter een ovaal masker. Middenpaneel toont een monster niet-zintuiglijk geometrisch object, terwijl rechterpaneel de Fourier-fase gerandomiseerde versie van objecten toont, gebruikt als controleprikkels.,
gegevensverzameling en-analyse
beeldvorming werd uitgevoerd met behulp van een 3-T MR-Headscanner (Siemens Allegra, Erlangen, Duitsland). Voor de functionele serie hebben we continu beelden verkregen (29 plakjes, 10° gekanteld ten opzichte van axiaal, T2* gewogen EPI-sequentie, TR = 2000 ms; te = 30 ms; flip angle = 90°; 64 × 64 matrices; in-plane resolutie: 3 mm × 3 mm; plakdikte: 3 mm). Hoge-resolutie sagittale T1-gewogen beelden werden verkregen met behulp van een magnetisatie EPI sequentie (MP-RAGE; TR = 2250 ms; TE = 2.,6 ms; 1 mm isotropische voxelgrootte) om een 3D structurele scan te verkrijgen (voor details, zie Nagy et al., 2011).
functionele beelden werden gecorrigeerd voor acquisitievertraging, opnieuw uitgelijnd, genormaliseerd naar de MNI-ruimte, opnieuw bemonsterd tot 2 mm × 2 mm × 2 mm resolutie en ruimtelijk gladgestreken met een Gaussiaanse kernel van 8 mm FWHM (SPM8, Welcome Department of Imaging Neuroscience, London, UK; voor details van data-analyse, zie Nagy et al., 2011).
VOI selectie
eerst werden volumes van Interesses (VOI) geselecteerd op basis van activiteit en anatomische beperkingen (inclusief maskering voor relevante hersengebieden)., Gezicht-selectieve gebieden werden gedefinieerd als een gebied met een grotere activering voor gezichten in vergelijking met Fourier ruisbeelden en objecten. FFA werd gedefinieerd binnen de laterale spoelvormige Gyrus, terwijl OFA binnen de IOG. LO werd gedefinieerd vanuit het Object > Fourier ruis en gezichtsbeelden contrast, binnen de middelste occipitale Gyrus. VOI-selectie was gebaseerd op de T-contrasten aangepast met F-contrast (p < 0,005 niet gecorrigeerd met een minimale clustergrootte van 15 voxels)., Voi ‘ s waren bolvormig met een straal van 4 mm rond de piekactivering (voor individuele coördinaten, zie tabel A1 in aanhangsel). De variantie verklaard door de eerste eigenvariate van de vetgedrukte signalen was allemaal boven 79%. In de huidige DCM-analyse werden alleen de rechterhersenhelft-gebieden gebruikt, aangezien verschillende studies wijzen op de dominante rol van dit halfrond in gezichtswaarneming (Michel et al., 1989; Sergent et al., 1992).,
effectieve Connectiviteitsanalyse
effectieve connectiviteit werd getest door DCM-10, geà mplementeerd in SPM8 toolbox (Wellcome Department of Imaging Neuroscience, London, UK), uitgevoerd onder Matlab R2008a (The MathWorks, Natick, MA, USA). Modellen van DCM worden gedefinieerd met endogene verbindingen, die de koppeling tussen hersengebieden (matrix A), modulerende verbindingen (matrix B), en drijvende input (matrix C) vertegenwoordigen. Hier in de a-matrix hebben we de verbindingen gedefinieerd tussen de face-selective regio ‘ s (FFA en OFA) en LO., Beelden van gezichten en objecten dienden als drijvende input (matrix C) en bij deze analyse stap hebben we geen modulerende effecten toegepast op de verbindingen.
Modelschatting was gericht op het maximaliseren van de negatieve vrije-energieschattingen van de modellen (F) voor een gegeven dataset (Friston et al., 2003). Deze methode zorgt ervoor dat het model fit gebruikt de parameters op een spaarzame manier (Ewbank et al., 2011). De geschatte modellen werden vergeleken, gebaseerd op het model bewijsmateriaal p(y|m), dat is de waarschijnlijkheid p van het verkrijgen van waargenomen gegevens y gegeven door een bepaald model m (Friston et al., 2003; Stephan et al., 2009)., In deze studie passen we de negatieve vrije-energie benadering (variationele vrije-energie) toe op het log bewijs (MacKay, 2003; Friston et al., 2007). Bayesian Model selectie (BMS) werd uitgevoerd op zowel de random (RFX) en fixed (FFX) effect ontwerpen (Stephan et al., 2009). BMS RFX is beter bestand tegen uitschieters dan FFX en het gaat er niet van uit dat hetzelfde model de functie voor elke deelnemer zou verklaren (Stephan et al., 2009). Met andere woorden RFX is minder gevoelig voor ruis., In de RFX-benadering is de output van de analyse de overschrijdingskans van de modelruimte, wat de mate is waarin een model de gemeten gegevens waarschijnlijker verklaart dan andere modellen. De andere output van de RFX-analyse is de verwachte posterieure waarschijnlijkheid, die de waarschijnlijkheid weerspiegelt dat een model de waargenomen gegevens genereerde, waardoor verschillende distributies voor verschillende modellen mogelijk zijn. Beide parameterwaarden worden verminderd door de verbreding van de modelruimte (d.w.z.,, door het aantal modellen te verhogen), daarom gedragen ze zich relatief en modellen met gedeelde functies en ongeloofwaardige modellen kunnen de output van de analyse verstoren. Daarom, in aanvulling op de directe vergelijking van de 28 gemaakte modellen hebben we het model ruimte verdeeld in families, met vergelijkbare connectiviteit patronen, met behulp van de methoden van Penny et al. (2010).
Aangezien verschillende eerdere DCM-studies wijzen op de nauwe bidirectionele verbinding tussen FFA en OFA (Ishai, 2008; Gschwind et al.,, 2012) in onze Analyse waren deze twee gebieden altijd met elkaar verbonden en LO was er op alle biologisch plausibele wijze mee verbonden. De 28 relevante modellen werden verdeeld in drie modelfamilies op basis van structurele verschillen (Penny et al., 2010; Ewbank et al., 2011). Familie 1 bevat modellen met lineaire verbindingen tussen de drie gebieden, ervan uitgaande dat informatie van de LO naar de FFA via de OFA stroomt. Familie 2 bevat modellen met een driehoekige structuur waarbij LO input direct naar de FFA stuurt en de OFA ook direct gekoppeld is aan de FFA., Familie 3 bevat modellen, waarin de drie gebieden met elkaar verbonden zijn, uitgaande van een cirkelvormige informatiestroom (Figuur 2). Om het aantal modellen in deze stap van de analyse te beperken, moduleerde de input uitsluitend de activiteit van hun entry areas. De drie families werden vergeleken door een willekeurig ontwerp BMS.
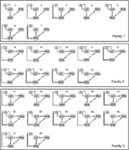
Figuur 2. De 28 geanalyseerde modellen. Zwarte lijnen markeren de verdeling tussen de drie families, met verschillende a matrix structuren. Voor details, zie Sectie ” materialen en methoden.,”
ten tweede werden de modellen van de winner-familie verder uitgewerkt door elk plausibel model met modulatoire verbindingen te creëren, waarbij drie beperkingen werden toegepast. Ten eerste moduleren zowel gezichten als objecten ten minste één Inter-Areale verbinding. Ten tweede, in het geval van bidirectionele koppelingen hebben de modulerende inputs een effect op beide richtingen. Ten derde, als face een directe input geeft in OFA dan gaan we ervan uit dat het altijd ook de OFA–FFA verbinding moduleert (zie tabel A2 in bijlage). Deze modellen werden ingevoerd in een tweede familie-wise random BMS analyse., Tot slot werden leden van de winner subfamilie opgenomen in een derde BMS om een enkel model te vinden met de hoogste overschrijdingswaarschijnlijkheid.
resultaten
Bayesiaanse modelselectie werd gebruikt om te bepalen welke modelfamilie de gemeten gegevens het beste verklaart. Zoals onze resultaten tonen de derde familie overtroffen de andere twee, met een overschrijdingskans van 0,995 vergeleken met de eerste familie 0,00 en de tweede familie 0,004 (figuren 3A,B)., De winner model familie (familie 3; Figuur 2, onder) bevat 12 modellen, met verbindingen tussen de LO en FFA, OFA en FFA, en LO en OFA ook, maar verschillend in de richting van de verbindingen en in de plaats van de input aan het netwerk.

Figuur 3. Resultaten van BMS RFX op het niveau van families. (A) de verwachte waarschijnlijkheden van familiegebaseerde vergelijking worden getoond, met de gezamenlijke overschrijdingswaarschijnlijkheden (B).,
als tweede stap werden alle mogelijke moduleringsmodellen ontworpen voor de 12 modellen van familie 3 (Zie tabel A2 in bijlage; materialen en methoden voor details). Dit leidde tot 122 modellen, die werden ingevoerd in de BMS familie-wise willekeurige analyse, met behulp van 12 subfamilies. Zoals uit de figuren 4A en B te zien is,heeft model-subfamilie 4 de andere subfamilies met een overschrijdingskans van 0,79 overtroffen. Als derde stap werden de modellen binnen de winner sub-family 4 ingevoerd in een willekeurig effect BMS. Figuur 5 toont de 18 geteste varianten van Model 20., Het model met de hoogste overschrijdingswaarschijnlijkheid (p = 0,75) was model 4 (Zie figuur 6). Dit betekent dat het winner-model bidirectionele verbindingen bevat tussen alle gebieden en gezicht-en objectingangen, verrassend genoeg, komen beide in de LO. Bovendien hebben vlakken een modulerend effect op de verbinding tussen LO en FFA, terwijl objecten de verbinding tussen LO en OFA moduleren.

Figuur 4. Resultaten van BMS RFX op het niveau van subfamilies., (A) de verwachte waarschijnlijkheden van familiegebaseerde vergelijking worden getoond, met de gezamenlijke overschrijdingswaarschijnlijkheden (B).

Figuur 5. Modellen van subfamilie 4. Alle modellen hebben dezelfde DCM.Een structuur (identiek aan model 20 in familie 3), DCM.B structuur verschilt van model tot model. Gestippelde pijlen gesymboliseerd gezicht modulerende effect, terwijl gestippelde vierkante pijlen toont object modulatie.
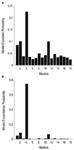
Figuur 6., Resultaten van BMS binnen subfamilie 4. (A) de verwachte waarschijnlijkheden van familiegebaseerde vergelijking worden getoond, met de gezamenlijke overschrijdingswaarschijnlijkheden (B).
Voor het analyseren van parameterschattingen van het winner-model over de groep proefpersonen werd een random effect benadering gebruikt. Alle subject-specific maximum a posteriori (MAP) schattingen werden ingevoerd in een t-test voor single means en getest tegen 0 (Stephan et al., 2010; Desseilles et al., 2011). De met een sterretje geïndexeerde resultaten op Figuur 7 verschilden aanzienlijk van 0 (p < 0.,05).
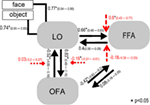
Figuur 7. De structuur van het winner model. Eenvoudige lijnen betekenen het object en het gezicht input stimuli naar het systeem (DCM.C). Zwarte pijlen tonen interregionale verbindingen (DCM.A) terwijl de rode pijlen staan voor de modulerende verbindingen (DCM.B): gezichtsmodulatie wordt aangegeven met gestreepte pijlen, terwijl objectmodulatie wordt aangegeven met vierkante gestreepte pijlen. Gemiddelden op groepsniveau van KAARTSCHATTINGEN en betrouwbaarheidsintervallen van 95% worden geïllustreerd., De gemiddelden werden getest tegen 0 en significante resultaten worden aangegeven met * als p < 0,05.,
discussie
Het belangrijkste resultaat van de huidige effectieve connectiviteitsstudie suggereert dat (a) LO rechtstreeks verbonden is met het OFA–FFA face-processing systeem via bidirectionele verbindingen naar beide gebieden; (b) niet-face en face ingangen worden gemengd op het niveau van occipito-temporele gebieden en het systeem binnenkomen via LO; (c) face input heeft een modulerend effect op LO en FFA verbinding, terwijl object input moduleert de lo en ofa verbinding aanzienlijk.
de rol van LO in objectperceptie is bekend uit eerdere studies (Malach et al.,, 1995; Grill-Spector et al., 1998a, b, 2000; Lerner et al., 2001, 2008). Hoewel eerdere studies meestal een verhoogde activiteit aan het licht brachten voor complexe objecten, evenals voor gezichten in LO, wordt het gebied meestal geassocieerd met objecten en wordt relatief minder belang toegekend aan zijn rol in de verwerking van de pijler. In dit onderzoek plaatste een effectieve connectiviteitsanalyse de LO in het kernnetwerk van gezichtsperceptie. De directe link tussen OFA en FFA is eerder zowel functioneel als anatomisch bewezen (Gschwind et al., 2012)., Aangezien er momenteel echter geen gegevens beschikbaar zijn over de rol van LO in dit systeem, hebben we het op verschillende plausibele manieren gekoppeld aan de andere twee regio ‘ s.
het eerste random BMS toonde aan dat LO in de winner modelfamilie verbonden is met zowel FFA als OFA en dat de verbindingen bidirectioneel zijn. Daarom wordt benadrukt dat LO een directe structurele verbinding met FFA kan hebben., Bij het modelleren van alle mogelijke modulerende effecten vonden we dat een subfamilie van modellen won, waarbij zowel object-als gezichtsinputs het systeem binnenkomen via de LO, stel dat de LO een algemene en belangrijke inputregio rol speelt. Sinds eerdere functionele connectiviteitsstudies zijn alle begonnen met de analyse van het face-processing netwerk op het niveau van IOG, overeenkomend met Ofa (Fairhall and Ishai, 2007; Ishai, 2008; Cohen Kadosh et al., 2011; Dima et al., 2011; Foley et al., 2012) het is niet verwonderlijk dat ze de belangrijke rol van LO over het hoofd zagen., Echter, gezichten zijn eigenlijk een aparte categorie van visuele objecten, wat suggereert dat neuronen gevoelig voor objecten en vormen zouden moeten worden geactiveerd, ten minste tot op zekere hoogte, door gezichten ook. Inderdaad, eencellige studies van niet-menselijke primaten suggereren dat de inferieure-temporale cortex, de voorgestelde homoloog van menselijke LO in de makaken hersenen (Denys et al., 2004; Sawamura et al., 2006) heeft ook neuronen die reageren op gezichten (Perrett et al., 1982, 1985; Desimone et al., 1984; Hasselmo et al., 1989; Young and Yamane, 1992; Sugase et al., 1999)., De intieme verbinding van LO met OFA en FFA, voorgesteld door de huidige studie zou het feit kunnen onderstrepen dat gezichten en objecten niet volledig afzonderlijk in de ventrale visuele route worden verwerkt, een conclusie die ook door recente functionele beeldvormingsgegevens wordt ondersteund (Rossion et al., 2012). Het modulerende effect van de pijlerinput op de LO-FFA-verbinding suggereert dat LO een rol moet spelen in de pijlerbewerking, hoogstwaarschijnlijk gekoppeld aan de vroegere, structurele verwerking van pijlers, een taak die eerder voornamelijk aan Ofa (Rotshtein et al., 2005; Fox et al., 2009)., Ten slotte tonen de parameters op groepsniveau van het winner-model aan dat de link tussen de LO, OFA en FFA voornamelijk hiërarchisch is en dat de feedbackverbindingen van FFA naar OFA en LO een zwakkere rol spelen in het systeem, althans tijdens de toegepaste fixatietaak.
concluderend, door het modelleren van de effectieve connectiviteit tussen Gezicht-relevante gebieden en LO suggereren we dat LO een belangrijke rol speelt in de verwerking van gezichten.,
belangenconflict verklaring
De auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
Dankbetuigingen
Dit werk werd ondersteund door de Deutsche Forschungsgemeinschaft (KO 3918/1-1) en de Universiteit van Regensburg. Wij danken Ingo Keck en Michael Schmitgen voor de discussie.
Campbell, R. (2011). Speechreading en het Bruce-Young model van gezichtsherkenning: vroege bevindingen en recente ontwikkelingen. Br. J. Psychol. 102, 704–710.,
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Fairhall, S. L., and Ishai, A. (2007). Effectieve connectiviteit binnen het gedistribueerde corticale netwerk voor gezichtswaarneming. Cereb. Cortex 17, 2400-2406.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Hoffman, E. A., and Haxby, J. V. (2000). Duidelijke representaties van oogopslag en identiteit in het gedistribueerde menselijke neurale systeem voor gezichtswaarneming. Nat. Neurowetenschappen. 3, 80–84.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Kourtzi, Z.,, and Kanwisher, N. (2001). Representatie van waargenomen objectvorm door het menselijk laterale occipitale complex. Wetenschap 293, 1506-1509.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
MacKay, D. J. C. (2003). Informatie theorie, gevolgtrekking, en leren algoritmen. Cambridge: Cambridge University Press.
Nasanen, R. (1999). Ruimtelijke frequentiebandbreedte die wordt gebruikt bij de herkenning van gezichtsopnamen. Vision Res. 39, 3824-3833.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Rossion, B., Schiltz, C., and Crommelinck, M., (2003b). De functioneel gedefinieerde rechter occipitale en spoelvormige “gezichtsgebieden” onderscheiden roman van visueel bekende gezichten. Neuroimage 19, 877-883.
CrossRef Full Text
Sawamura, H., Orban, G. A., and Vogels, R. (2006). Selectiviteit van neuronale adaptatie komt niet overeen met respons selectiviteit: een eencellige studie van het fMRI adaptatieparadigma. Neuron 49, 307-318.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
















Geef een reactie