Bevezető
Chloroplasts jelentenek strukturális jellemzője, hogy a növényi sejtekben, valamint támogatja a növény túlélési keresztül az elsődleges anyagcsere, valamint a magas szintű funkciók (Kirk Tilney-Bassett, 1978; Márna, 1988; López-Juez, valamint Pyke, 2005). A növényi vegetatív növekedés során a levélsejtek méretük és alakjuk tekintetében rendkívül homogén kloroplaszt populációt tartalmaznak., A sejtenkénti kloroplasztok számát A már létező organellák bináris hasadásával érik el, és a sejtosztódás során két lánysejtre osztódnak (Birky, 1983; Possingham and Lawrence, 1983). Így a kloroplasztszám szabályozása egy adott sejttípusban döntő fontosságú a kloroplasztok sejtfunkciójához és genetikai örökléséhez.
a növényi sejtekben a kloroplasztszám meghatározásának természetének vizsgálata érdekében a reprezentatív Fajok levélmezofillsejtjei jelentős szerepet játszottak ., Ezek a sejtek fiziológiailag fontosak a fotoszintézis szempontjából, és nagyfokú szerkezeti és funkcionális homogenitást mutatnak. Az izolált szövetek és sejtek korai szisztematikus megfigyelési elemzése (Boasson and Laetsch, 1969; Possingham and Saurer, 1969; Boffey et al., 1979; Lamppa et al.,, 1980; Thomas and Rose, 1983; Pyke and Leech, 1991) sok hasznos információt szolgáltatott a kloroplasztszám meghatározásáról, beleértve azt az elképzelést, hogy a kloroplasztokat (plasztidokat) nem szintetizálják de novo, hanem osztással replikálják, valamint azt a megfigyelést, hogy a levél mezofill kloroplaszt száma érzékeny a különböző környezeti és növényi endogén tényezőkre. Ez utóbbi tekintetében a spenótban a fény pozitív hatással van a kloroplaszt osztódására a leveles lemezkultúra során, összehasonlítva a homályos vagy sötét állapotokkal (Possingham and Lawrence, 1983)., A búza első leveleiben a sejtek térfogata pozitív korrelációban van a kloroplaszt proliferációval (Ellis and Leech, 1985; Pyke and pióca, 1987). Az Arabidopsis-ban a genetikai háttér befolyásolja a kloroplaszt proliferációját; az első levelekben a sejtenkénti átlagos kloroplasztszám 121 A Landsberg erecta (Ler) ekotípusban és 83 a Wassilewskija (Ws) ekotípusban (Pyke and Leech, 1994; Pyke et al., 1994)., Levél mesophyll sejtek is hozzájárult ahhoz, hogy megértsük a genetikai ellenőrzése chloroplast osztály; például a szűrés mutánsok csökkent a chloroplast elterjedése, valamint jellemző gén funkciók részt chloroplast osztály tártak fel, több mint 20 gének kódoló chloroplast osztály gépek, berendezések, vagy chloroplast szabályozó tényezők (Gao, valamint Gao, 2011; Miyagishima et al., 2011; Basak and Møller, 2013; Osteryoung and Pyke, 2014; Li et al., 2017).
ezzel szemben a kloroplasztok nem mezofillsejtekben történő replikációjára vonatkozó vizsgálatok (pl.,, járda sejtek levél epidermisz; Itoh et al., 2018) szűkös. A közelmúltban a kloroplaszt osztódás szabályozásáról számoltak be, hogy különböznek a levélszövetek között (Fujiwara et al., 2018; Itoh et al., 2018), bár a részletes mechanizmus ismeretlen marad. Továbbá, míg az analízis a szuszpenzió-tenyésztett BY-2 sejtek és levél mezofill protoplasztok dohány és lőni apikális merisztém és levél primordiális sejtek Arabidopsis (Nebenführ et al., 2000; Sheahan et al.,, 2004; Seguí-Simarro és Staehelin, 2009) jelentős betekintést nyújtott, hogy a kloroplaszt (plastid) particionálás hogyan szabályozható a növényekben, még mindig nem tisztázott. Így, annak ellenére, hogy jelentős erőfeszítéseket, alapvető kérdése, hogy chloroplast kutatási marad, például, (i) hogy chloroplast szám per sejt összehangolt a növényi szövetekben, valamint (ii) hogyan chloroplast particionálás szabályozza a sejtosztódást.,
Történelem Kutatás Őr Sejt Chloroplast Száma
Stomatal GCs a lő bőr általában tartalmaznak chloroplasts kontroll -, gáz közötti levél mesophyll a légkör (Sachs, 1875; Taiz et al., 2015; lásd az 1a. ábrát). A levelek GC-kloroplasztszámának első vizsgálatát több mint egy évszázaddal ezelőtt végezték el a természetesen termesztett Drosera növényekben (Macfarlane, 1898)., Ez a tanulmány kimutatta, hogy, mint más növényi sejtek szerkezeti jellemzők, GC chloroplast száma cellánként egy vélelmezett hibrid származik, egy kereszt között Harmatfű filiformis, valamint Harmatfű intermedia volt köztes között a két faj, utalva arra, hogy a GC chloroplast száma lehetne használni, hogy meghatározzák a genetikai összetétele a növény. Ezt követően fontos megfigyelésekről számoltak be a GC kloroplasztszám különbségeiről a növényfajok között (Sakisaka, 1929), valamint az Eperfa (Morus spp.,; Hamada és Baba, 1930) és több Brassica faj Érett leveleiben (Iura, 1934). Ezenkívül az autopoliploid cukorrépa (béta vulgaris) növények elemzése kimutatta, hogy a levelekben lévő GC kloroplasztszám pozitívan korrelál a növények nukleáris ploidi szintjével (Mochizuki and Sueoka, 1955). Ezután mélyrehatóbb és átfogóbb elemzéseket végeztek különböző növényminták felhasználásával a kloroplasztszám és a sztóma méretének a ploidi szinttel való összefüggésének vizsgálatára (pl. Frandsen, 1968)., Ezekben az elemzésekben gyakran alkalmazták a sztóma (GC pár) szintjén a kloroplaszt számlálást, amely kizárta a párosított GCs (pl. Mochizuki és Sueoka, 1955; Frandsen, 1968) közötti elfogult kloroplaszt-Eloszlás hatását, feltárva, hogy a levelekben vagy sziklevelek átlagos GC-kloroplasztszáma körülbelül 80 fajban, változatban vagy hibridben 2,8-40,0 volt diploidokban (2×) és 5,0-73,5 között tetraploidokban (4×). Ezenkívül a teljes genom párhuzamos eseményei növényekben (azaz 1× 2×, 2× 4× stb.) okozott kb. 1.,A GC kloroplasztszám 7-szeres növekedése nagy hűséggel (áttekintve Butterfass, 1973). Ezek az eredmények arra ösztönözték a ploidia szintjének vizsgálatát különböző szövetekben és növényekben, amelyeket szövetkultúra, keresztezés vagy természetes termesztés útján, vegyi anyagokkal (pl. kolchicin) vagy sugárkezelésekkel (pl. Jacobs and Yoder, 1989; Singsit and Veilleux, 1991; Qin and Rotino, 1995) kombinálva nyertek., Míg a GC chloroplast száma tanulmányozták stomatal biológia (Lawson, 2009), citológia, hogy megértsük, chloroplast szorzás (Butterfass, 1979; lásd alább), jobbára szolgált, mint egy megbízható, kényelmes jelölő kimutatására hibridek, fajok, illetve változatokat, valamint a becslés ploidy szinten a cél a növényi szövetekben.
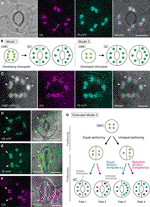
1.ábra egyszerűsített modellek és mikroszkópos bizonyítékok a sztomatális védősejtekben (GCs) található kloroplasztszám szabályozására., (A) Egy tipikus sztóma (GC pár) a abaxial felhám az Osztályába levéllemez kifejező stoma-célzott fúziós a tranzit peptid (TP) a cián fluoreszcens fehérje (KHP; TP-CFP). B) a Kloroplasztszám meghatározásának két modellje a GCs-ben, amely magában foglalja a kloroplaszt particionálását (1.modell) vagy a kloroplaszt proliferációját és a particionálást (2. modell) a GC fejlesztése során az anyasejtekből (GMCs). C) A zöld fluoreszcens fehérjével (GFP; FtsZ1-GFP) kondenzált Arabidopsis levélnyél adaxiális epidermiszében lévő GC pár., (D–F) az Arabidopsis levélpenge abaxiális epidermiszében lévő GC Párok (D, E) vagy anélkül (F) A sárga fluoreszkáló fehérjéhez (YFP; TP-YFP) olvasztott TP kifejezéssel. F) klorofill-autofluoreszcenciát (Chl) használtak kloroplaszt markerként., (G) a Kiterjesztett modell, 2, képviselő bevonása az egyenlő, illetve egyenlő chloroplast particionálás következő GMC osztály, valamint az azt követő osztály GC chloroplasts egyenlő (kék vonal), vagy szelektív (piros vonal) osztály kompetencia, amely azt eredményezné, hogy négy típusú chloroplast szám meghatározása (Sors 1-4) során késő stomatal fejlesztési Osztályába levelek. (A, C–F) az Epifluoreszcens mikroszkópiát Olympus Ix71 invertált mikroszkóppal végezték a korábban leírt növényi anyagok felhasználásával (Fujiwara et al., 2017, Fujiwara et al., 2018)., A klorofill, a CFP, a GFP és az YFP fluoreszcenciajelei álszínűek bíbor, Cián, Zöld (csak egyesített képen), illetve zöld színben. A panelek jelzései a következők: nyílhegy, az FtsZ1 gyűrű; nyíl, nagyított GC kloroplaszt; szaggatott vonal, sejt alak. Skála bar = 10 µm.
A Levélvédő sejtek hasznossága a Kloroplasztszám-Szabályozás
levél mezofillsejtek elemzéséhez már régóta elsődleges modellként alkalmazzák a kloroplasztszám elemzéséhez., Bár előnyeik vannak a környezeti feltételek kloroplaszt osztódásra gyakorolt hatásának tanulmányozására (pl. a fény-citokinin jelátvitel; Boasson and Laetsch, 1969; Possingham and Lawrence, 1983; Okazaki et al., 2009; Chiang et al., 2012), bizonyos tekintetben korlátozottak. Először is, a levél mesophyll sejtek mérete és alakja változó, és mélyen eloszlik a levélen belül, ami megnehezíti az ép szövetek manipulálását., Másodszor, a levél mezofill kloroplaszt proliferációjának a környezeti stresszre és a növény növekedési körülményeire való érzékenysége megakadályozhatja a vizsgálatok közötti megbízható összehasonlítást. Az Arabidopsis Columbia (Col) ecotípus sejtenkénti mezofill-kloroplaszt számát különböző módon jelentették 76 (Kinsman and Pyke, 1998), 80-100 (Stokes et al., 2000), 70 (Tirlapur and König, 2001), 41 (Yoder et al., 2007), 40-60 (Okazaki et al., 2009) és 30-40 (Kawade et al., 2013)., Ezenkívül szinte lehetetlen felmérni a kloroplaszt particionálásnak a végső kloroplasztszámhoz való hozzájárulását mezofillsejtenként a levélfejlődés során, bár ezt úgy gondolják, hogy a sejtosztódás és a kloroplasztosztódás aránya közötti egyensúly határozza meg. A kloroplasztszám-szabályozás mechanizmusának feltárásához vegetatív levélsejtekben olyan modellrendszerre van szükség, amely legyőzi a fenti kérdéseket.
Stomatal GCs (lásd az 1a.ábrát) a kloroplasztszám-szabályozás mechanizmusának megértésére szolgáló Modellrendszer jellemzőit mutatja., A GC-k mérete és alakja rendkívül egyenletes a szöveten belül, és szétszórt, de sűrű eloszlásuk a hajtások legkülső rétegében megkönnyíti fény-és fluoreszcens mikroszkóppal történő kimutatásukat. GCs is származik protodermális sejtek a hajtás apikális merisztémában vagy embrionális epidermális sejtek, és azok fejlődési szekvenciáját merisztemoidok (egy sztómatikus prekurzor merisztematikus aktivitás) és őr anyasejtek (GMC; prekurzor GC Párok) állapítják meg részletesen (Zhao and Sack, 1999; Nadeau and Sack, 2002; Kalve et al., 2014)., A késői sztomatális fejlődés a szimmetrikus GMC-divízió egyetlen körét foglalja magában, amely lehetővé teszi a kloroplaszt Eloszlás és a citokinezis előtti és utáni particionálás értékelését. A gyakorlati kísérletek szempontjából a levél GCs mikroszkópiára alkalmas. Korábban kimutatták, hogy a Sinapis alba levélgcseiben található sejtenkénti kloroplasztszámot kevésbé befolyásolta a különböző fényviszonyok, mint a levél mezofillsejtekben (vad és Farkas, 1980)., Továbbá, a különbség a GC kloroplaszt száma levél levélnyél viszonylag kisebb között a három Arabidopsis ökotípusok Col, Ler, és Ws (Fujiwara et al., 2018). Az Arabidopsis leaf GCS (Melaragno et al., 1993), amely biztosítja a kloroplasztszám-adatok értelmezését a sejtek 2C szintjén., Ezek a jelentések együttesen azt sugallják, hogy a leaf GCs potenciálisan kiváló modell a kloroplasztszám dinamikájának szisztematikus elemzéséhez egy adott sejtvonalban.
Osztályába Levél Őr Sejtek, mint egy Modell a Tanulás Ellenőrzése Chloroplast Száma
a történelem GC chloroplast kutatás, chloroplast számítva a stomatal (GC pár) szinten szolgált egy hasonlóan fontos szerepet játszik annak meghatározásában, a chloroplast száma, mint számítva az egyedi GC szinten. Mindkét módszer ugyanazt az átlagos kloroplasztszámot állítja elő (Butterfass, 1973)., Ha a kloroplaszt-Eloszlás változása a párosított GCs-ben és annak mögöttes mechanizmusa fókuszpont alatt áll, a kloroplasztok részletes információja az egyes sejtek szintjén, azaz méretük, alakjuk és intracelluláris lokalizációjuk elengedhetetlen. A kloroplaszt (plastid) proliferációját a GMC–GC differenciálás során korábban több növényfajban vizsgálták (Butterfass, 1973, Butterfass 1979)., Ezek a vizsgálatok javasolt két modell meghatározására a terminál chloroplast száma a GCs a különböző növényfajok (1B Ábra): egy (modell 1; cukorrépa) magában csak chloroplast particionálás a GMC osztály, a másik magában foglalja nem csak chloroplast particionálással, de egy is chloroplast elterjedése során GC fejlesztés.
a molekuláris genetika, a genomika, a sejt képalkotás és más interdiszciplináris elemzések korában számos lehetőség van a kloroplaszt particionáló mechanizmus további jellemzésére., Arabidopsis leaf GCS lehet az egyik legjobb modell rendszerek erre a célra. Számos tanulmány vizsgálta a GC kloroplaszt számát az Arabidopsis leveleiben vagy szikleveleiben (Hoffmann, 1968; Pyke and Leech, 1994; Pyke et al., 1994; Robertson et al., 1995; Keech et al., 2007; Chen et al., 2009; Yu et al., 2009; Higaki et al., 2012; Fujiwara et al., 2018). Ezek a GC-k szerény számú kloroplasztot mutatnak, átlagosan 3, 5-5, 5 között. A mai napig egyetlen tanulmány sem vizsgálta a kloroplaszt (plastid) számának változásait a sztomatális fejlődés során., Azonban mikroszkopikus bizonyítékok a gyomorfejlődési elemzésekből (pl. Zhao and Sack, 1999; Hachez et al., 2011) és előzetes megfigyeléseink azt mutatják, hogy a GMC-k kisebb számban tartalmazhatnak fejlődő kloroplasztokat, mint a GCs-k, és hogy a kloroplaszt proliferációja előfordulhat a GC differenciálódása során. Ennek tesztelésére a kloroplaszt osztású gép GCS-ben történő képződését a zöld fluoreszcens fehérjével (FtsZ1-GFP) (Fujiwara et al., 2008)., A transzgenikus vonalat, amely egyidejűleg tranzitpeptid (TP)-fuzionált CFP-t, valamint FtsZ1-GFP-t fejezett ki a stroma, illetve az FtsZ1 gyűrű megjelenítésére, epifluoreszcens mikroszkópiával (Fujiwara et al., 2017). Bővülő levélnyéleket (a 4 hetes palánták ötödik leveleit) alkalmaztak. Ennek eredményeképpen szimmetrikusan összehúzódó kloroplasztokkal rendelkező GCs-ket észleltek (1C ábra). Ezek a kloroplasztok képezték az FtsZ1 gyűrűt, egy kloroplaszt osztógyűrűt a belső borítékmembrán stromális felületén levél mezofillben és járda sejtekben (Vitha et al., 2001; Fujiwara et al.,, 2008), az egyenlítői szűkület helyén. Az Arabidopsis leaf development (Donnelly et al., 1999; Andriankaja et al., 2012), a kloroplasztok elosztását a levél terjeszkedésének késői, valamint korai szakaszában észlelték. Így a 2. modell valószínűleg a legjobban illeszkedik az Arabidopsis leaf GCS-hez.
Kapcsolat Chloroplast Elterjedése, Valamint Terjeszkedés Osztályába Levél Őr Sejtek
Továbbá, egy váratlan fenotípus GC chloroplast morphogenesis volt megfigyelhető érett GCs (Adatok 1D, E)., Amikor epidermális hámlasztja teljesen kiterjesztett levelek (harmadik-negyedik levél pengék 4 hetes palánták) egy TP-olvasztott sárga fluoreszkáló fehérje (YFP) vonal mikroszkóposan jellemezhető (FL6-5 vonal; Fujiwara et al., 2018), néhány sztóma egyenlőtlen kloroplaszt eloszlási mintákat mutatott GC párokban, míg a legtöbb levél sztóma egyenlő vagy hasonló kloroplaszt eloszlási mintákat mutatott (Robertson et al., 1995; Fujiwara et al., 2018)., A sztóma GC-párján belül a GC-ben kisebb számú kloroplasztot tartalmazó kloroplasztok mérete nagyobb volt, mint a pár többi GCs-jében, amely nagyobb számú kloroplasztot tartalmaz (1D, E ábra). Ily módon a GCs valószínűleg a sejtenkénti teljes kloroplaszt térfogatot állandó szinten tartja a sejtnövekedés során. A megnagyobbodott kloroplasztok a terminális fenotípust képviselték, és már nem oszthatók szét kiterjesztett levelekben. Ezeket az eredményeket számos független kísérletben megerősítették, függetlenül a TP-vel fuzionált fluoreszcens fehérje kifejeződésétől a stroma címkézéséhez (1F ábra).,
ezt a GC kloroplaszt fenotípust a kloroplaszt expanzió kompenzációs mechanizmusaként értelmezik, amelyet jól dokumentáltak a kloroplaszt divízió szabályozásában hibás leveles mezofillsejtekben (Pyke and Leech, 1994; Pyke et al., 1994). A mai napig csak egy tanulmány (Ellis and Leech, 1985) számolt be negatív korrelációról a kloroplasztszám és a kloroplasztméret között a búza leveles mezofillsejtjeiben, mivel számos tanulmány pozitív korrelációról számolt be a sejtek térfogata és a kloroplasztszám között a normál levél mezofillsejtekben (Leech and Pyke, 1988; Pyke, 1997)., Míg a GC kloroplasztszám egyensúlyhiányai alacsony frekvencián fordulnak elő (Fujiwara et al., 2018), a kloroplaszt heterogenitása a GC párokban azt jelzi, hogy az egyenlőtlen kloroplaszt particionálás differenciál kloroplaszt növekedést válthat ki az Arabidopsis vad típusú levélsejtjei között, a szimmetrikus sejtosztódás ellenére.
a kloroplaszt kompenzációs hatása a GCs-ben kevésbé szigorú lehet, mint a levél mezofill sejtekben., GCs lehet, hogy képes ellenállni szűkössége vagy teljes elvesztése teljes kloroplaszt térfogat sejtenként súlyosan károsodott kloroplaszt osztódás mutánsok, mint például az Arabidopsis arc6 és atminE1 és paradicsom suffulta, mivel sok mutáns GCs mutatott csökkentett kloroplaszt száma és megnagyobbodott kloroplaszt mérete hasonlóan a levél mezofill sejtek (Robertson et al., 1995; Forth and Pyke, 2006; Chen et al., 2009; Fujiwara et al., 2018)., A késői kloroplaszt osztódás mutánsában, az arc5-ben a GC kloroplasztszám csökkenése nem társult a kloroplaszt méretének jelentős növekedésével, ellentétben a levél mezofillsejtekkel (Pyke and Leech, 1994). A kloroplaszt-expanzió alacsonyabb foka a GCs-ben, mint a mezofill sejtekben (Pyke and Leech, 1994; Barton et al., 2016), valamint a kloroplaszt expanziójának változása a GCs között, a teljes kloroplaszt térfogat / GC ilyen széles megengedett tartományát alapozhatja meg., Ezenkívül a kloroplaszt osztódásának időzítése a GMC-GC differenciálódás során jelentősen befolyásolhatja a terminális GC kloroplaszt fenotípust. Bár a probléma megoldásához további részletes jellemzésre van szükség, valószínűnek tűnik, hogy az Arabidopsis leaf GCs egy olyan rendszert képvisel, amely megvizsgálja a kloroplasztszám-szabályozás kihasználatlan aspektusait a növényi sejtekben.,
az Arabidopsis Levélvédő sejtekben a Kloroplasztszám meghatározására szolgáló munkamodell
a fentiek alapján egy működő modellt (egy kiterjesztett 2.modellt) javasolunk a Kloroplasztszám GCS-ben történő elemzéséhez (1g ábra). A GC-n belüli végső kloroplasztszámot a GMC-divízióban történő kloroplaszt-particionálás és a GCs-ben a kloroplaszt proliferáció határozza meg. A GMC felosztása során a kloroplasztok egyenlő vagy egyenlőtlen felosztáson eshetnek át. A kloroplaszt proliferációja során a GC kloroplasztok egyenlő (kék vonal) vagy szelektív (magenta vonal) osztódási képességgel szaporodnak., Például, ha az egyenlően felosztott kloroplasztok egyenértékű osztási kompetenciával rendelkeznek, akkor a GC párban egyenlő kloroplaszt számok fordulnak elő (1.sors). Ha az egyenlőtlenül felosztott kloroplasztok egyenértékű osztódási képességgel rendelkeznek, a kloroplasztok ugyanolyan sebességgel növekednek a GC páron belül (3.sors). Ha a GCS-ben szelektív kloroplasztosztódás következik be, a GC-párban a kloroplasztszám egyenlege megváltozik a GMC-felosztás után (2.és 4. sors)., Jelenleg nehéz megtalálni a “szelektív kloroplasztosztódás” támogatását, de ha az 1.és a 4. sors ténylegesen túlsúlyban van a GCs-ben, akkor rendelkezhetnek olyan mechanizmussal, amely szabályozza a sejtenkénti teljes kloroplaszt térfogatot, mint a levél mezofill sejtekben. A modell két kérdést vet fel: (i) a GC-kloroplasztok megfelelően vannak felosztva leánysejtekbe, és hogyan osztódnak szét? És ii. a GC-kloroplasztok megosztási kompetenciája koordináltan szabályozott?,
ami az i. kérdést illeti, hogy a kloroplaszt öröklés a többszörös kloroplasztok véletlenszerű eloszlásával történik-e a citoplazmában vagy pozitív kloroplaszt-particionáló mechanizmus(ok) révén, régóta aggodalomra ad okot (Butterfass, 1969; Birky, 1983; Hennis and Birky, 1984; Nebenführ, 2007; Sheahan et al., 2016). Érdekes módon az Arabidopsis arc6-ban a levél vagy a sziklevelek GCs-jének nulla-három kloroplasztja van, a kloroplaszt-hiányos GCs-ben pedig a nem fotoszintetikus plasztidok továbbra is léteznek hólyagos vagy hosszúkás formákban (Robertson et al., 1995; Chen et al., 2009; Fujiwara et al., 2018)., Az arc6-ban nem találtak plasztidoktól mentes GCs-t, és erre a kloroplaszt osztóberendezés (Vitha et al., 2003). Ennek megfelelően fontos lesz megvizsgálni az arc6-kloroplasztok replikációját és morfológiáját a sztomatális lineage vizsgálatokban. Arabidopsis mutáns kutatás is ad egy másik nyom ebben a kérdésben., Az a megfigyelés, hogy a gyűrött levél (crl) mutánsban a sziklevelek GCS 18% – a nem tartalmaz plasztid struktúrákat a citoplazmában, míg a levél mezofill sejtek 100% – a Tartalmaz egy-négy megnagyobbodott kloroplasztot (Asano et al., 2004; Chen et al., 2009), nagyon fontos. A CRL egy kloroplaszt külső borítékú fehérje, ismeretlen funkcióval. A CRL fehérje funkció megértése betekintést nyújthat a kloroplaszt particionálás mechanizmusába(mechanizmusaiba)., Az Arabidopsis arc mutánsok és más transzgenikus vonalak levélmezofill sejtjeiben a kloroplaszt proliferáció és particionálás elemzése elősegítette a nem-mezofill plasztidok proliferációjának és particionálásának kutatását. Hasonlóképpen, a GCs-ben kapott eredmények átvihetők más cellarendszerekre.
záró Megjegyzés
a GC modell számos lehetőséget kínál a kloroplaszt biológia fejlesztésére. Például, míg a cytoskeletalis rendszerekről ismert, hogy szabályozzák a kloroplaszt morfológiáját, mozgását és particionálását (Sheahan et al.,, 2016; Wada, 2016; Erickson and Schattat, 2018), Az egyes szabályozó gének szerepe a kloroplaszt proliferációban és a növényekben való felosztásban kevés figyelmet kapott. Másrészt, ha lehetővé válik a GC kloroplaszt számának vagy morfológiájának károsítása különböző kísérleti stratégiák révén, új betekintést lehet nyújtani a sztomatális Vonalú sejtek kloroplaszt morfogenezisének molekuláris szabályozásába. Továbbá, a sztomatális fejlődés során a kloroplaszt szám kvantitatív elemzésével együtt, a matematikai modellezés új utat kínálhat ezeknek a vizsgálatoknak., Ez a tanulmány bemutatja a GC kloroplasztszám szabályozásának jelenlegi ismereteit, és kiemeli az Arabidopsis leaf GCS potenciális hasznosságát a kloroplaszt proliferáció és a particionálás megértésében.
az adatok rendelkezésre állásáról szóló nyilatkozat
A tanulmány összes adatkészlete szerepel a cikkben/ kiegészítő anyagban.
szerzői hozzájárulások
az MF megfogalmazta a tanulmányt és megírta a kéziratot. MF, valamint végzett kísérletek. AS és RI végezte az elemzéseket. Minden szerző elolvasta és jóváhagyta a végleges kéziratot.,
finanszírozás
ezt a munkát a japán oktatási, kulturális, tudományos és Technológiai Minisztérium támogatta KAKENHI alatt (19k05831-MF támogatási szám és 18K06314-RI).
összeférhetetlenség
a szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.
elismerések
a szerzők köszönetet mondanak Dr. Nobuyuki Kanzawa – nak (Sophia Egyetem) a munkával kapcsolatos tanácsokért.
Birky, C. W., Jr. (1983)., A citoplazmatikus organellák felosztása a sejtosztódásban. Int. Cytol Tiszteletes. 15, 49-86.
Google Scholar
butter churn, T. (1969). A haploid swedenklee (Trifolium hybridum L.) zárósejtes anyasejtjeinek mitózisában a plastid Eloszlás. Planta 84., 230-234. doi: 10.1007/BF00388108 P>PubMed Abstract | CrossRef teljes szöveg/Google Scholar
butter churn, T. (1973). A plasztidosztódás ellenőrzése nukleáris DNS-mennyiséggel. Protoplazma 76, 167-195. doi: 10.,1007/BF01280696
CrossRef Full Text | Google Scholar
Qin, X., Rotino, G. L. (1995). Chloroplast number in guard cells as ploidy indicator of in vitro-grown androgenic pepper plantlets. Plant Cell Tissue Organ Cult. 41, 145–149. doi: 10.1007/BF00051583
CrossRef Full Text | Google Scholar
Sachs, J. (1875). Textbook of Botany. Oxford: Clarendon Press.
Google Scholar
Thomas, M. R., Rose, R. J. (1983)., A plastid száma és a plastid szerkezeti változásai a dohány mezofill protoplaszt tenyészetével és a növény regenerációjával összefüggésben. Planta 158, 329-338. doi: 10.1007/BF00397335
PubMed Abstract | CrossRef teljes szöveg/Google Scholar
Tirlapur, U. K., König, K. (2001). Femtosecond közel infravörös lézerek, mint egy új eszköz a nem invazív valós idejű nagy felbontású time-lapse képalkotó kloroplaszt Osztás élő köteg köpeny sejtek Arabidopsis. Planta 214, 1-10. doi: 10.,1007 / s004250100597
PubMed Abstract / CrossRef teljes szöveg / Google Scholar
Wild, A., Wolf, G. (1980). A különböző fényintenzitások hatása a sztóma gyakoriságára és méretére, a sejtek méretére, a kloroplasztok számára, méretére és klorofilltartalmára a mezofillben és a Védősejtekben a Sinapis alba elsődleges leveleinek ontogenitása során. Z. Pflanzenphysiol. Bd. 97, 325–342. doi: 10.1016 / S0044-328x (80)80006-7
CrossRef teljes szöveg | Google Scholar
















Vélemény, hozzászólás?