Úvod
Chloroplasty, které představují strukturální funkce rostlinných buněk a podpůrných rostlinných přežití prostřednictvím jejich primární metabolismus a funkce na vysoké úrovni (Kirk a Tilney-Bassett, 1978; Parmice, 1988; Lopez-Juez a Štítu, 2005). Během vegetativního růstu rostlin obsahují listové buňky vysoce homogenní populaci chloroplastů vzhledem k velikosti a tvaru., Počet chloroplastů na buňku je dosažen binárním štěpením již existujících organel a rozdělením na dvě dceřiné buňky během buněčného dělení (Birky, 1983; Possingham a Lawrence, 1983). Regulace chloroplastového čísla v daném buněčném typu je tedy zásadní pro buněčnou funkci a genetickou dědičnost chloroplastů.
pro zkoumání povahy stanovení počtu chloroplastů v rostlinných buňkách hrály hlavní roli listové mezofylové buňky reprezentativních druhů ., Tyto buňky jsou fyziologicky důležité pro fotosyntézu a vykazují vysoký stupeň strukturální a funkční homogenity. Počátkem systematického pozorování, analýzy izolovaných tkání a buněk (Boasson a Laetsch, 1969; Possingham a Saurer, 1969; Boffey et al., 1979; Lamppa et al.,, 1980; Thomas a Rose, 1983; Pyke a Leech, 1991), za předpokladu, mnoho užitečných informací na chloroplast počet stanovení, včetně představa, že chloroplasty (plastidy) nejsou syntetizovány de novo, ale replikaci rozdělení a zjištění, že list mesophyll chloroplastu číslo je citlivý na různé oblasti životního prostředí a rostlina-endogenní faktory. Pokud jde o posledně jmenované, ve špenátu má světlo pozitivní vliv na rozdělení chloroplastů během kultivace listových disků ve srovnání s tlumenými nebo tmavými podmínkami (Possingham a Lawrence, 1983)., V prvních listech pšenice je objem buněk pozitivně korelován s proliferací chloroplastů (Ellis a pijavice, 1985; Pyke a pijavice, 1987). V Arabidopsis, genetické pozadí ovlivňuje chloroplastu šíření; průměrný chloroplast, počtu na buňku, a to v první listy je 121 v Landsberg erecta (Ler) ekotypu a 83 v Wassilewskija (Ws) ekotypu (Pyke a Pijavice, 1994; Pyke et al., 1994)., List mesophyll buňky mají také přispěl k pochopení genetické kontroly chloroplastu divize; například, screening mutanti postižené v chloroplastu šíření a charakterizaci funkce genů, zapojených v chloroplastu divize odhalily více než 20 genů kódujících chloroplastu divize strojní součásti nebo chloroplast regulační faktory (Gao a Gao, 2011; Miyagishima et al., 2011; Basak and Møller, 2013; Osteryoung and Pyke, 2014; Li et al., 2017).
naproti tomu studie replikace chloroplastů v nemesofylových buňkách (např.,, dlažební buňky v listové epidermis; Itoh et al. , 2018) jsou vzácné. Nedávno bylo hlášeno, že regulace chloroplastového dělení se liší mezi listovými tkáněmi (Fujiwara et al., 2018; Itoh et al., 2018), ačkoli podrobný mechanismus zůstává neznámý. Navíc, zatímco analýzy suspenzí kultivovaných buněk-2 a listových mezofylových protoplastů v tabáku a střílet apikální meristem a listové prvotní buňky v Arabidopsis(Nebenführ et al ., 2000; Sheahan et al.,, 2004; Seguí-Simarro a Staehelin, 2009) poskytly významné poznatky, jak je rozdělení chloroplastů (plastid) regulováno v rostlinách, je stále nejasné. A tak, i přes značné úsilí, základní otázky v chloroplastu výzkumu zůstat, jako (já), jak je chloroplast, počtu na buňku, a koordinovány v rostlinných tkáních, a (ii) jak je chloroplast rozdělení regulovány na buněčné dělení.,
Historie Výzkumu na Stráži Buňky Chloroplastu Číslo
Stomatal GCs v natáčení epidermis obvykle obsahují chloroplasty a řízení výměny plynů mezi listem mesophyll a atmosféru (Sachs, 1875; Taiz et al., 2015; viz obrázek 1a). První zkoumání počtu chloroplastů GC v listech bylo provedeno před více než stoletím v přirozeně pěstovaných rostlinách Drosera (Macfarlane, 1898)., Tato studie prokázala, že, stejně jako ostatní rostliny a buňky strukturální rysy, GC chloroplast, počtu na buňku, a to v předpokládaných hybridů odvozených z křížení mezi Drosera filiformis a Drosera intermedia byla prostředníkem mezi dvěma druhy, což znamená, že GC chloroplastu číslo může být použit k určení genetické make-up rostliny. Důležitá pozorování byla následně hlášena o rozdílech v počtu chloroplastů GC u druhů rostlin (Sakisaka, 1929) a relativně stabilním počtu chloroplastů v GCs v listové epidermis Moruše (Morus spp.,; Hamada a Baba, 1930) a ve zralých listech několika druhů Brassica (Iura, 1934). Kromě toho, analýza autopolyploid cukrové řepy (Beta vulgaris) rostliny bylo zjištěno, že GC chloroplastu číslo v listech pozitivně koreluje s jadernou úroveň ploidie rostlin (Mochizuki a Sueoka, 1955). Podrobnější a komplexnější analýzy byly poté provedeny pomocí různých vzorků rostlin, aby se prozkoumal vztah počtu chloroplastů a velikosti stomatu s hladinou ploidy (např., V těchto analýzách, počítání chloroplastů v stomatal (GC pár) úroveň byla často přijata, což vyloučen vliv neobjektivní chloroplastu distribuce mezi spárovanými GCs (např. Mochizuki a Sueoka, 1955; Frandsen, 1968), odhaluje, že průměrná GC chloroplastu číslo listů nebo děložními lístky v přibližně 80 druhů, variant, nebo hybridů se pohybovala od 2,8 až 40.0 v neznámý (2×) a 5.0 až 73,5 v tetraploids (4×). Kromě toho dochází k duplikaci celého genomu v rostlinách (tj. 1× až 2×, 2× až 4× atd.) způsobil přibližně 1.,7-násobné zvýšení počtu chloroplastů GC s vysokou věrností (přezkoumáno v Butterfassu, 1973). Tyto výsledky vybízejí vyšetřování ploidie úrovni v různých tkáních a rostliny získané přes tkáňové kultury, přejezd, nebo přírodní pěstování, v kombinaci s chemickou látkou (např. kolchicin) nebo ozařováním (např. Jacobs a Yoder, 1989; Singsit a Veilleux, 1991; Qin a Rotino, 1995)., Zatímco GC chloroplastu počet byl zkoumán v stomatal biologie (Lawson, 2009) a cytologie pochopit, chloroplast, násobení (Butterfass, 1979; viz níže), má do značné míry sloužil jako spolehlivý a vhodný marker pro detekci hybridů, druhů a variant a pro odhad úrovně ploidie cílových rostlinných tkání.
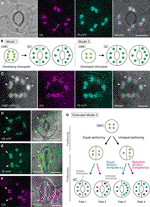
Obrázek 1 Zjednodušené modely a mikroskopie důkazy pro kontrolu chloroplastu číslo v stomatal stráž buněk (GCs)., A) typická stomie (dvojice GC) v abaxiální epidermis listu Arabidopsis vyjadřující stomicky cílenou fúzi tranzitního peptidu (TP) s azurovým fluorescenčním proteinem (CFP; TP-CFP). (B) Dva modely chloroplastu počet stanovení na Gc, zahrnující buď chloroplastu rozdělení (model 1) nebo oba chloroplast, proliferaci a dělení (model 2) během GC vývoj od strážce matka buněk (GMCs). C) dvojice GC v adaxiální epidermis řapíku Arabidopsis Leaf exprimující TP-CFP a FtsZ1 fúzovaná se zeleným fluorescenčním proteinem (GFP; FtsZ1-GFP)., (D–F) páry GC v abaxiální epidermis listu Arabidopsis S (D, E) nebo bez (F) exprese TP fúzované se žlutým fluorescenčním proteinem (YFP; TP-YFP). F) chlorofyl autofluorescence (Chl) byl použit jako chloroplastový marker., (G) Rozšířený model 2, což představuje zapojení rovné a nerovné dělení chloroplastu následující GMC divize a následné rozdělení GC chloroplasty se rovná (modrá čára) nebo selektivní (červená čára), rozdělení kompetencí, což by mělo za následek čtyři typy chloroplastů počet stanovení (Osudy 1-4) během pozdní stomatal vývoje Arabidopsis listy. (A, C-F) epifluorescenční mikroskopie byla provedena s obráceným mikroskopem Olympus IX71 za použití rostlinných materiálů, jak bylo popsáno výše (Fujiwara et al., 2017, Fujiwara et al., 2018)., Fluorescenční signály chlorofylu, CFP, GFP a YFP jsou pseudobarevné v purpurové, azurové, zelené (pouze ve sloučeném obrazu)a zelené. Indikace v panelech jsou následující: Šipka, prstenec FtsZ1; šipka, zvětšený chloroplast GC; přerušovaná čára, tvar buňky. Měřítko = 10 µm .
Utility Listu Stráž Buněk Pro Analýzu Chloroplastu Kontrolní Číslo
List mesophyll cells již dlouho zaměstnán jako primární model pro analýzu chloroplastu číslo., Zatímco oni mají své výhody pro studium vlivu podmínek prostředí na chloroplast divize (např. světlo-hodnotami signalizace; Boasson a Laetsch, 1969; Possingham a Lawrence, 1983; Okazaki et al., 2009; Chiang et al., 2012), jsou v některých ohledech omezené. Za prvé, listové mezofylové buňky se liší velikostí a tvarem a jsou distribuovány hluboko uvnitř listu, což ztěžuje manipulaci s neporušenými tkáněmi., Za druhé, náchylnost proliferace listového mezofylu chloroplastu k environmentálnímu stresu a podmínkám růstu rostlin může zabránit spolehlivému srovnání mezi studiemi. V ekotypu Arabidopsis Columbia (Col) bylo hlášeno číslo chloroplastu listového mezofylu na buňku různě jako 76 (Kinsman a Pyke, 1998), 80-100 (Stokes et al., 2000), 70 (Tirlapur a König, 2001), 41 (Yoder et al., 2007), 40-60 (Okazaki et al., 2009) a 30-40 (Kawade et al., 2013)., Kromě toho, že je téměř nemožné posoudit přínos chloroplastu dělení na konečné chloroplastu číslo za mesophyll buňku v listu vývoje, i když to je si myslel, že stanoví rovnováhu mezi mírou buněčného dělení a rychlost dělení chloroplastu. K odhalení mechanismu kontroly počtu chloroplastů ve vegetativních listových buňkách je vyžadován modelový systém, který překonává výše uvedené problémy.
stomatální GCs (viz obrázek 1a) vykazují vlastnosti modelového systému pro pochopení mechanismu kontroly počtu chloroplastů., GCs jsou ve tkáni vysoce jednotné velikosti a tvaru a jejich rozptýlené, ale husté rozložení v nejvzdálenější vrstvě výhonků usnadňuje jejich detekci světelnou a fluorescenční mikroskopií. GCs jsou také odvozeny od protodermal buněk v shoot apical meristem nebo z embryonálních buněk epidermis, a jejich vývojové sekvence prostřednictvím meristemoids (stomatal předchůdce s meristematic činnost) a stráž matka buněk (GMCs; prekurzor GC párů) je stanovena v detailu (Zhao a Sack, 1999; Nadeau a Sack, 2002; Kalve et al., 2014)., Pozdní stomatální vývoj zahrnuje jedno kolo symetrického dělení GMC, které umožňuje posouzení distribuce chloroplastů a rozdělení před a po cytokinezi. Z pohledu praktických experimentů jsou leaf GCs vhodné pro mikroskopii. To bylo již dříve prokázáno, že chloroplast číslo na buňku v listu GCs z Sinapsis alba byla méně ovlivněna při různých světelných podmínkách, než v listu mesophyll cells (Divoké a Wolf, 1980)., Kromě toho je rozdíl v počtu chloroplastů GC v listových řapících relativně malý mezi třemi ekotypy Arabidopsis Col, Ler a Ws (Fujiwara et al., 2018). Endoreduplikace, která ovlivňuje vývoj listového mezofylu, dlažby a trichomových buněk, navíc nebyla zjištěna u GCs listů Arabidopsis (Melaragno et al ., 1993), což by zajistilo interpretace údajů o počtu chloroplastů na úrovni buněk 2C., Spolu, tyto zprávy naznačují, že list GCs jsou potenciálně vynikající model pro systematickou analýzu chloroplastu číslo dynamika v konkrétní buněčné linie.
Arabidopsis List Stráž Buňky jako Model pro Studium Kontrolu Chloroplastu Číslo
V historii GC chloroplastu výzkumu, počítání chloroplastů v stomatal (GC pár) úroveň sloužil stejně důležitou roli při určování chloroplastu číslo jako počítání na individuální GC úrovni. Obě metody produkují stejné průměrné číslo chloroplastu (Butterfass, 1973)., Pokud je změna distribuce chloroplastů ve spárovaných GCs a jejím základním mechanismu předmětem zaměření, jsou nezbytné podrobné informace o chloroplastech na úrovni jednotlivých buněk, tj. jejich velikost, tvar a intracelulární lokalizace. Proliferace chloroplastů (plastid) během diferenciace GMC–GC byla dříve zkoumána u několika druhů rostlin (Butterfass, 1973, Butterfass 1979)., Tyto studie navrhuje dva modely pro určení terminálu chloroplastu číslo v GCs v různých rostlinných druhů (Obrázek 1B): (model 1; cukrové řepy) zahrnuje pouze chloroplastu rozdělení na GMC divize, a jiné zahrnuje nejen dělení chloroplastu, ale také chloroplast šíření během GC rozvoj.
v době molekulární genetiky, genomiky, zobrazování buněk a dalších interdisciplinárních analýz existuje mnoho možností pro další charakterizaci mechanismu dělení chloroplastů., Arabidopsis leaf GCS může být jedním z nejlepších modelových systémů pro tento účel. Několik studií zkoumalo číslo chloroplastu GC v listech nebo kotyledonech Arabidopsis (Hoffmann, 1968; Pyke a pijavice, 1994; Pyke et al., 1994; Robertson et al., 1995; Keech et al., 2007; Chen et al., 2009; Yu et al., 2009; Higaki et al., 2012; Fujiwara et al., 2018). Tyto GCs vykazují mírný počet chloroplastů, v průměru od 3,5 do 5,5. K dnešnímu dni žádné studie nezkoumaly změny v počtu chloroplastů (plastid) během stomatálního vývoje., Nicméně, mikroskopické důkazy z analýzy vývoje stomatální (např., Zhao a Sack, 1999; Hachez et al., 2011) a naše předběžná pozorování ukazují, že GMCs může obsahovat menší počet rozvojových chloroplasty než GCs a že chloroplastu šíření může dojít během GC diferenciace. Pro tento test, vznik chloroplastu divize strojů v GCs byla sledována pomocí sondy FtsZ1 taveného pro zelený fluorescenční protein (FtsZ1-GFP) (Fujiwara et al., 2008)., Transgenní linie, současně vyjadřuje tranzitního peptidu (TP)-tavený SRP a FtsZ1-GFP pro vizualizaci stroma a FtsZ1 prsten, respektive, byl vyšetřen epifluorescence mikroskopie (Fujiwara et al., 2017). Byly použity rozšiřující se listové řapíky (páté listy 4týdenních sazenic). V důsledku toho byly detekovány GCs se symetricky zúženými chloroplasty (obrázek 1C). Tyto chloroplasty tvoří FtsZ1 prsten, chloroplast divize prsten na stromální povrch na vnitřní obálce membrány v listu mesophyll a chodníku buněk (Vitha et al., 2001; Fujiwara et al.,, 2008), v místě rovníkového zúžení. V souladu se stomatálním vzorováním ve vývoji listů Arabidopsis (Donnelly et al., 1999; Andriankaja et al., 2012), dělící chloroplasty byly detekovány v pozdním, stejně jako časném stádiu expanze listů. Model 2 je tedy s největší pravděpodobností nejvhodnější pro listy Arabidopsis GCS.
vztah mezi proliferací chloroplastů a expanzí v buňkách ochrany listů Arabidopsis
u zralých GCs byl navíc pozorován neočekávaný fenotyp morfogeneze chloroplastu GC (obrázky 1D, E)., Když epidermální peeling z plně rozvinutými listy (třetí–čtvrtý list čepele 4-týden-staré semenáčky) od TP-tavený žlutý fluorescenční protein (YFP) linie byly mikroskopicky charakterizována (FL6-5 řádek; Fujiwara et al., 2018), některé průduchy ukázal nerovné distribuce chloroplastů vzory v GC párů, zatímco většina listových průduchů ukázal stejný nebo podobný chloroplastu distribuce (Robertson et al., 1995; Fujiwara et al., 2018)., V páru GC stomie byla velikost chloroplastů v GC obsahující menší počet chloroplastů větší než u ostatních GC v páru obsahujícím větší počet chloroplastů (obrázky 1D, E). Tímto způsobem GCs pravděpodobně udržuje celkový objem chloroplastu na buňku na konstantní úrovni během růstu buněk. Zvětšené chloroplasty představovaly terminální fenotyp a již se nemohly dělit v expandovaných listech. Tyto výsledky byly potvrzeny v několika nezávislých experimentech, bez ohledu na expresi fluorescenčního proteinu taveného TP pro označování stroma (obrázek 1F).,
Tento GC chloroplastu fenotyp je interpretován jako kompenzační mechanismus pro chloroplastu expanzi, což bylo dobře zdokumentované v listu mesophyll buňky defektní v kontrole dělení chloroplastu (Pyke a Pijavice, 1994; Pyke et al., 1994). K dnešnímu dni, pouze jedna studie (Ellis a Pijavice, 1985) uvádí negativní korelaci mezi chloroplast, počet a velikost chloroplastu v listu mesophyll cells pšenice, vzhledem k tomu, že mnoho studií uvádí pozitivní korelaci mezi objem buněk a chloroplastů číslo v normálním list mesophyll cells (Pijavice a Štítu, 1988; Pyke, 1997)., Zatímco nerovnováha v počtu chloroplastů GC se vyskytuje při nízké frekvenci(Fujiwara et al ., 2018), heterogenita chloroplastů v párech GC naznačuje, že nerovnoměrné rozdělení chloroplastů by mohlo vyvolat diferenciální růst chloroplastů mezi listovými buňkami divokého typu v Arabidopsis, navzdory symetrickému dělení buněk.
účinek kompenzace chloroplastů v GCs může být méně přísný než v buňkách mezofylu listů., GCs by mohl být schopen vydržet nedostatek nebo úplná ztráta celkový objem chloroplastu na buňku poruchou chloroplastu divize mutanty, jako v Arabidopsis arc6 a atminE1 a rajče suffulta, vzhledem k tomu, že mnoho mutant GCs vykazovali sníženou chloroplastu číslo a rozšířené chloroplastu velikost podobně jako list mesophyll cells (Robertson et al., 1995; Forth a Pyke, 2006; Chen et al., 2009; Fujiwara et al., 2018)., V pozdním mutantu chloroplastového dělení, arc5, nebylo snížení počtu chloroplastů GC spojeno s významným zvýšením velikosti chloroplastu, na rozdíl od buněk mezofylu listů (Pyke a pijavice, 1994). Nižší stupeň chloroplastové expanze v GCs než v mezofylových buňkách (Pyke a pijavice, 1994; Barton et al., 2016) a rozdíly v expanzi chloroplastů mezi GCs mohou být základem tak širokého přípustného rozsahu celkového objemu chloroplastů na GC., Kromě toho může načasování dělení chloroplastů během diferenciace GMC–GC významně ovlivnit terminální fenotyp chloroplastu GC. Přestože je k řešení tohoto problému zapotřebí další podrobná charakterizace, zdá se pravděpodobné, že GC listů Arabidopsis představují systém pro zkoumání nevyužitých aspektů kontroly počtu chloroplastů v rostlinných buňkách.,
Pracovní Model Pro Chloroplastu Počet Stanovení v Arabidopsis List Stráž Buňky
Na základě výše uvedeného navrhujeme, pracovní model (rozšířený model 2) pro analýzu chloroplastu číslo v GCs (Obrázek 1G). Konečné číslo chloroplastu na GC je určeno rozdělením chloroplastů při dělení GMC a proliferací chloroplastů v GCs. Během dělení GMC mohou chloroplasty procházet rovným nebo nerovným rozdělením. Během proliferace chloroplastů se chloroplasty GC proliferují buď se stejnou (modrou čarou) nebo selektivní (purpurovou čarou) divizí., Například, pokud stejně rozdělené chloroplasty mají ekvivalentní kompetenci rozdělení, stejná čísla chloroplastů se vyskytnou v páru GC (osud 1). Pokud nerovnoměrně rozdělené chloroplasty mají ekvivalentní divizní kompetenci, chloroplasty se zvýší stejným tempem v páru GC (osud 3). Pokud dojde k selektivnímu dělení chloroplastů v GCs, změní se rovnováha chloroplastového čísla v páru GC po rozdělení GMC (osudy 2 a 4)., V současné době je obtížné najít podporu pro „selektivní dělení chloroplastu,“ ale pokud Osud 1 a 4 ve skutečnosti převládají v GCs, pak oni by mohli mít mechanismus, který řídí celkový objem chloroplastu v buňce, stejně jako v listu mesophyll cells. Model vyvolává dva problémy: (i) jsou chloroplasty GC správně rozděleny do dceřiných buněk a jak se rozdělují? A (ii)je kompetence GC chloroplastů koordinovaně regulována?,
O problému (i), zda chloroplastu dědictví dochází tím, že náhodné rozdělení více chloroplastů v cytoplazmě nebo pozitivní dělení chloroplastu mechanismus(y) byl dlouholetým zájmem (Butterfass, 1969; Birky, 1983; Hennis a Birky, 1984; Nebenführ, 2007; Sheahan et al., 2016). Kupodivu, v Arabidopsis arc6, list nebo cotyledon GCs mít nula až tři chloroplasty, a v chloroplastu-nedostatečné GCs, non-fotosyntetizující plastidy stále existují v vezikulární do podlouhlé formy (Robertson et al., 1995; Chen et al., 2009; Fujiwara et al., 2018)., Ne GCs bez plastidů per se, byly nalezeny v arc6, a žádné vysvětlení byla nastávající, a to navzdory narušení chloroplastu divize přístroje (Vitha et al., 2003). Proto bude důležité zkoumat replikaci a morfologii chloroplastů arc6 ve studiích stomatální linie. Arabidopsis mutantní výzkum může také dát další vodítko pro tento problém., Zjištění, že 18% z děložních GCs v zmačkaný list (crl) mutant neobsahují žádné plastidic struktur v cytoplazmě, zatímco 100% list mesophyll buňky obsahují jeden až čtyři rozšířené chloroplasty (Asano et al., 2004; Chen et al., 2009), má velký význam. CRL je chloroplastový vnější obalový protein s neznámou funkcí. Pochopení funkce proteinu CRL může poskytnout pohled na mechanismus (y) rozdělení chloroplastů., Analýza proliferace a dělení chloroplastů v listových mezofylových buňkách v mutantech Arabidopsis arc a dalších transgenních liniích podpořila výzkum proliferace a dělení nemetofylových plastidů. Podobně mohou být výsledky získané v GCs přenositelné do jiných buněčných systémů.
závěrečná poznámka
model GC otevírá mnoho vyhlídek na vývoj chloroplastové biologie. Například, zatímco cytoskeletální systémy jsou známé regulovat chloroplast morfologii, pohyb ,a dělení (Sheahan et al.,, 2016; Wada, 2016; Erickson a Schattat, 2018), úloha každého regulačního genu při proliferaci a dělení chloroplastů v rostlinách získala malou pozornost. Na druhou stranu, jakmile to bude možné narušit GC chloroplastu číslo nebo morfologie pomocí různých experimentálních strategií, nové pohledy na molekulární kontrolu morfogenezi chloroplastů v stomatal linie buněk mohou být poskytnuty. Navíc, ve spojení s kvantitativní analýzy chloroplastu číslo během stomatal rozvoj, matematické modelování může nabídnout nové avenue pro tyto výzkumy., Tento článek představuje současné znalosti o tom, jak je kontrolováno číslo chloroplastu GC, a zdůrazňuje potenciální užitečnost GC listů Arabidopsis pro pochopení proliferace a dělení chloroplastů.
Prohlášení o dostupnosti dat
všechny datové sady pro tuto studii jsou zahrnuty v článku / doplňkovém materiálu.
autorské příspěvky
MF koncipovalo studii a napsalo rukopis. MF a AS provedl experimenty. AS a RI provedli analýzy. Všichni autoři přečetli a schválili konečný rukopis.,
finanční Prostředky
Tato práce byla podpořena Ministerstvem Školství, Kultury, Vědy a Technologie Japonska pod KAKENHI (grant nos. 19K05831 na MF a 18K06314 na RI).
střet zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
poděkování
autoři děkují Dr. Nobuyuki Kanzawa (Sophia University) za radu ohledně práce.
Birky, C. W., Jr. (1983)., Rozdělení cytoplazmatických organel na buněčné dělení. Int. Rev. Cytol. 15, 49-86.
Google Scholar
Butter churn, T. (1969). Plastidová distribuce v mitóze uzavíracích buněčných mateřských buněk haploid swedenklee (Trifolium hybridum L.). Planta 84, 230-234. doi: 10.1007/BF00388108
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
máslo máselnice, T. (1973). Řízení plastidového dělení pomocí množství nukleární DNA. Protoplasma 76, 167-195. doi: 10.,1007/BF01280696
CrossRef Full Text | Google Scholar
Qin, X., Rotino, G. L. (1995). Chloroplast number in guard cells as ploidy indicator of in vitro-grown androgenic pepper plantlets. Plant Cell Tissue Organ Cult. 41, 145–149. doi: 10.1007/BF00051583
CrossRef Full Text | Google Scholar
Sachs, J. (1875). Textbook of Botany. Oxford: Clarendon Press.
Google Scholar
Thomas, M. R., Rose, R. J. (1983)., Plastidové číslo a plastidové strukturální změny spojené s tabákovou mezofylovou protoplastovou kulturou a regenerací rostlin. Planta 158, 329-338. doi: 10.1007/BF00397335
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Tirlapur, U. K., König, K. (2001). Femtosekundový blízké infračervené lasery jako román nástroj pro neinvazivní real-time high-resolution time-lapse imaging of chloroplast divize v obývacím bundle sheath cells of Arabidopsis. Planta 214, 1-10. doi: 10.,1007 / S004250100597
PubMed Abstract | CrossRef Full Text / Google Scholar
Wild, a., Wolf, G. (1980). Vliv různé světelné intenzity na frekvenci a velikost průduchů, velikost buněk, počet, velikost a obsah chlorofylu z chloroplastů v mesophyll a stráž buněk během ontogeneze primární listy Sinapsis alba. Z. Pflanzenfysiol. Bd. 97, 325–342. doi: 10.1016 / S0044-328X (80)80006-7
CrossRef Full Text / Google Scholar
















Napsat komentář