Johdanto
Kloroplastit ovat rakenteellinen ominaisuus kasvin soluja ja tukemaan kasvin selviytymisen kautta heidän ensisijainen aineenvaihduntaa ja korkean tason toimintoja (Kirk ja Tilney-Bassett, 1978; Keltti, 1988; López-Juez ja Piikkiin, 2005). Kasvien kasvullisen kasvun aikana lehtisoluissa on erittäin homogeeninen kloroplastipopulaatio koon ja muodon suhteen., Määrä viherhiukkasia per solu on saavuttaa binary fissio ennestään soluelimiin ja jakautuminen kahteen tytär soluja solunjakautumisen aikana (Birky, 1983; Possingham ja Lawrence, 1983). Näin ollen asetuksen kloroplastiin numero tietyn solun tyyppi on ratkaisevan tärkeää, jotta solujen toiminta ja geneettinen perintö viherhiukkasia.
tutkiakseen kloroplastiluvun määrityksen luonnetta kasvisoluissa edustavien lajien lehtimesofyllisoluilla on ollut suuri merkitys ., Nämä solut ovat fysiologisesti tärkeitä fotosynteesin kannalta ja niissä on paljon rakenteellista ja toiminnallista homogeenisuutta. Alussa systemaattinen havainnointi analyysit eristetty kudosten ja solujen (Boasson ja Laetsch, 1969; Possingham ja Saurer, 1969; Boffey et al., 1979; Lamppa ym.,, 1980; Thomas ja Nousi, 1983; Piikkiin ja Iilimato, 1991) saatiin paljon hyödyllistä tietoa kloroplastiin määrä päättäväisyyttä, mukaan lukien käsitys siitä, että viherhiukkasia (plastideissa) eivät ole synteettisesti de novo, mutta jäljitellä jako ja tarkkailu, että lehtiä mesophyll kloroplastiin numero on herkkä eri ympäristö-ja kasvi-endogeeniset tekijät. Osalta jälkimmäinen, pinaatti, valo on positiivinen vaikutus kloroplastiin jako aikana lehtiä levy kulttuuri verrattuna hämärässä tai pimeässä (Possingham ja Lawrence, 1983)., Ensimmäinen lehdet vehnä, solujen määrä korreloi positiivisesti kloroplastiin leviämisen (Ellis ja Iilimato, 1985; Piikkiin ja Iilimato, 1987). Arabidopsis, geneettinen tausta vaikuttaa kloroplastiin leviämisen; keskimääräinen kloroplastiin määrä per solu ensimmäiset lehdet on 121 Landsberg erecta (Ler) ekotyyppiä ja 83 Wassilewskija (Ws) ekotyyppiä (Pyke ja Iilimato, 1994; Piikkiin et al., 1994)., Lehtiä mesophyll solut ovat myös osaltaan ymmärrystä geneettinen ohjaus kloroplastiin jako; esimerkiksi seulonta mutantteja heikentynyt kloroplastiin leviämisen ja kuvaavat geenin toimintojen mukana kloroplastiin jako on paljasti yli 20 geenit koodaavat kloroplastiin jako koneiden osia tai kloroplastiin lainsäädännölliset tekijät (Gao Gao, 2011; Miyagishima et al., 2011; Basak and Møller, 2013; Osteryoung and Pyke, 2014; Li et al., 2017).
sen sijaan kloroplastien replikaatiota ei-mesofyllisoluissa koskevat tutkimukset (esim.,, pavement soluissa Leaf orvaskeden; Itoh et al., 2018) ovat niukat. Äskettäin asetuksen kloroplastiin jako on raportoitu vaihtelevan välillä lehtiä kudosten (Fujiwara et al., 2018; Itoh et al., 2018), vaikka yksityiskohtaista mekanismia ei vielä tunneta. Lisäksi, kun analyysit jousitus-viljellyt BY-2-solujen ja lehtiä mesophyll protoplasts tupakan ja ampua apical meristem ja lehtiä ensiarvoisen soluja Arabidopsis (Nebenführ et al., 2000; Sheahan et al.,, 2004; Seguí-Simarro ja Staehelin, 2009) ovat toimittaneet suuria oivalluksia, miten kloroplastiin (plastid) osiointi on säännelty kasvit on vielä epäselvä. Niinpä, vaikka paljon työtä, perustavanlaatuisia kysymyksiä kloroplastiin tutkimus on edelleen, kuten (i) miten on kloroplastiin numero solua kohti koordinoitua kasvi kudosten ja (ii) miten on kloroplastiin osiointi säädellä solujen jakautumista.,
Historia Tutkimusta Vartija Solu Kloroplastiin Numero
Ilmarakojen GCs ampua iho yleensä sisältävät viherhiukkasia ja hallita kaasun välistä lehtiä mesophyll ja ilmapiiri (Sachs, 1875; ta ’ izz et al., 2015; KS. Kuva 1a). Ensimmäinen tutkimus GC kloroplastiin numero lehdet tehtiin yli sata vuotta sitten luonnollisesti kasvanut Drosera kasvit (Macfarlane, 1898)., Tämä tutkimus osoitti, että, kuten muutkin kasvi, ja solun rakenteellisia ominaisuuksia, GC kloroplastiin numero solua kohti on oletettu hybridi peräisin risteytys Drosera filiformis ja Drosera intermedia oli välissä kaksi lajia, mikä tarkoittaa, että GC kloroplastiin numero voidaan määrittää geneettinen meikki kasvi. Tärkeitä havaintoja olivat myöhemmin kertoi eroja GC kloroplastiin numero yksi kasvilajeja (Sakisaka, 1929) ja suhteellisen vakaa kloroplastiin numero GCs-lehtiä iho mulberry (Morus spp.,; Hamada ja Baba, 1930) ja kypsä lehdet useita Brassica-lajien (Iura, 1934). Lisäksi analyysi autopolyploid sokerijuurikkaan (Beta vulgaris) kasvit paljasti, että GC kloroplastiin numero lehdet korreloi positiivisesti ydinvoimaan ploidy taso kasvit (Mochizuki ja Sueoka, 1955). Enemmän perusteellinen ja kattava analyysit olivat sitten tehdään käyttäen erilaisia kasvi näytteitä tutkia suhdetta kloroplastiin numero ja ilmarakojen kokoa ploidy taso (esim., Frandsen, 1968)., Näissä analyyseissä, kloroplastiin laskenta klo ilmarakojen (GC pari) taso oli usein hyväksyttiin, jonka ulkopuolelle vaikutus puolueellinen kloroplastiin jakautuminen pariksi GCs (esim., Mochizuki ja Sueoka, 1955; Frandsen, 1968), joka paljastaa, että keskimääräinen GC kloroplastiin numero lehtiä tai sirkkalehtien noin 80 lajia, muunnokset, tai hybridit vaihteli 2,8 40,0 vuonna totta on (2×) ja 5,0 73,5 vuonna tetraploids (4×). Lisäksi koko-genomin päällekkäisiä tapahtumia kasvit (eli 1×, 2×, 2×, 4×, jne.) aiheutti noin 1.,7-kertainen kasvu GC-klooriplastiluku korkea uskollisuus (arvosteltu Voirasvassa, 1973). Nämä tulokset kannusti tutkimuksia ploidy tasolla eri kudoksissa ja kasvien kautta saatuja kudosten kulttuuri -, risteämis -, tai luonnollinen viljely, yhdessä kemian (esim. kolkisiini) tai säteily hoitoja (esim, Jacobs ja Antao, 1989; Singsit ja Veilleux, 1991; Qin ja Rotino, 1995)., Kun GC kloroplastiin numero on tutkittu ilmarakojen biologia (Lawson, 2009), ja sytologia ymmärtää kloroplastiin kerto (Butterfass, 1979; ks. alla), se on pitkälti toiminut luotettava ja kätevä merkki havaitsemiseksi hybridit, laji, ja vaihtoehdot ja arvio ploidy tasoilla kohde kasvien kudoksia.
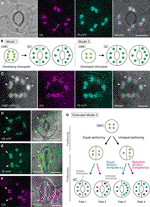
Kuva 1 Yksinkertaistettu malleja ja mikroskopia näyttöä valvonta kloroplastiin numero ilmarakojen vartija soluja (GCs)., (A) tyypillinen avanne (GC pari) vuonna abaxial orvaskeden Arabidopsis lehtiä terä ilmaista avanne-kohdennettu fuusio transit peptidi (TP), jossa on syaani fluoresoiva proteiini (YKP; TP-YKP). (B) Kaksi malleja kloroplastiin määrä päättäväisyyttä GCs, joissa joko kloroplastiin osiointi (malli 1) tai molemmat kloroplastiin leviämisen ja osiointi (malli 2) aikana GC kehittämiseen vartija äiti soluja (GMCs). (C) GC-pari adaxial orvaskeden Arabidopsis lehtien ruoti ilmaista TP-YKP: n ja FtsZ1 sulatetun vihreä fluoresoiva proteiini (GFP; FtsZ1-GFP)., (D–F) GC-parit abaxial orvaskeden Arabidopsis lehtiä terä (D, E) tai ilman (F) ilmaus TP sulatettu keltainen fluoresoiva proteiini (YFP; TP-YFP). (F) klorofyllin autofluoresenssia (Chl) käytettiin kloroplastimerkkinä., (G) Laajennettu malli 2, eli osallistumista, tasa-arvoisia ja epätasa-kloroplastiin osiointi seuraavat GMC jako ja myöhemmin jako GC viherhiukkasia yhtä (sininen viiva) tai valikoiva (punainen viiva) jako pätevyys, joka johtaisi neljä kloroplastiin numeron määritys (Fates 1-4) aikana myöhään ilmarakojen kehittäminen Arabidopsis lehdet. (A, C–F) Epifluorescence mikroskopointi suoritettiin Olympus IX71 käänteismikroskooppi käyttää kasvimateriaalit kuten aiemmin on kuvattu (Fujiwara et al., 2017, Fujiwara ym., 2018)., Fluoresenssi signaalit klorofylli, YKP, GFP, ja YFP ovat pseudo-värillinen magenta, syaani, vihreä (merged vain kuva), ja vihreä, vastaavasti. Paneelien merkinnät ovat seuraavat: nuolenkärki, ftsz1-rengas; nuoli, laajentunut GC-kloroplasti; murskattu viiva, solun muoto. Asteikko bar = 10 µm.
Utility Lehtiä Vartija Solujen Analysointiin Kloroplastiin Useita Ohjaus
Leaf mesophyll solut ovat jo pitkään olleet tuotto kuin ensisijainen malli analyysi kloroplastiin numero., Kun ne on etuja tutkimuksen ympäristöolosuhteiden vaikutus kloroplastiin jako (esim. valo-sytokiinin signalointi; Boasson ja Laetsch, 1969; Possingham ja Lawrence, 1983; Okazaki et al., 2009; Chiang et al., 2012), ne ovat tietyiltä osin rajallisia. Ensinnäkin, lehtiä mesophyll solut vaihtelevat kooltaan ja muodoltaan ja jaetaan syvällä leaf, mikä tekee siitä vaikea manipuloida ehjä kudoksiin., Toiseksi, herkkyys lehtiä mesophyll kloroplastiin leviämisen ympäristön stressiä ja kasvien kasvun edellytykset voivat estää luotettavia vertailuja tutkimukset. Arabidopsis Columbian (Col) ekotyypin mesofyllikloroplastilukua solua kohti on ilmoitettu vaihtelevasti 76 (Kinsman ja Pyke, 1998), 80-100 (Stokes et al., 2000), 70 (Tirlapur ja König, 2001), 41 (Yoder ym., 2007), 40-60 (Okazaki ym., 2009) ja 30-40 (Kawade et al., 2013)., Lisäksi, se on lähes mahdotonta arvioida panos kloroplastiin osiointi lopulliseen kloroplastiin määrä per mesophyll solu aikana lehtien kehitystä, vaikka tämä on ajateltu olevan määräytyy tasapaino määrä solunjakautumisen ja määrä kloroplastiin jako. Paljastaa mekanismi kloroplastiin numero-ohjaus kasvullisen lehtiä soluja, malli-järjestelmä, joka ratkaisee edellä mainitut kysymykset on tarpeen.
Ilmarakojen GCs (ks. Kuva 1A) näytteille ominaisuudet malli järjestelmän ymmärtämisen mekanismi kloroplastiin määrä control., GCs ovat erittäin tasakokoisia ja muodoltaan sisällä kudoksen, ja niiden hajallaan, mutta tiheä jakelu uloin kerros versoja helpottaa niiden havaitsemista valo-ja fluoresenssimikroskopia. GCs on myös johdettu protodermal soluissa ampua apical meristem tai alkion orvaskeden solujen, ja niiden kehityshäiriöitä järjestyksessä läpi meristemoids (a ilmarakojen esiaste kanssa meristematic toimintaa) ja vartija äiti soluja (GMCs; esiaste GC paria) on perustettu yksityiskohtaisesti (Zhao ja Säkki, 1999; Nadeau ja Säkki, 2002; Kalve et al., 2014)., Myöhään ilmarakojen kehitys sisältää yhden kierroksen symmetrinen GMC jako, joka mahdollistaa arvioinnin kloroplastiin jakelu ja jakaminen ennen ja jälkeen cytokinesis. Käytännön kokeiden näkökulmasta lehtien GCs soveltuu mikroskopiaan. Se oli aiemmin osoittanut, että kloroplastiin määrä per solu lehtiä GCs sekä Sinapis alba oli vähemmän vaikutusta eri valo-olosuhteissa kuin lehtien mesophyll solut (Wild ja Wolf, 1980)., Lisäksi ero GC kloroplastiin numero lehtiä petioles on suhteellisen vähäinen joukossa kolme Arabidopsis ekotyyppien Kol, Ler, ja Ws (Fujiwara et al., 2018). Lisäksi endoreduplication, joka vaikuttaa kehityksen lehtiä mesophyll, jalkakäytävä, ja trichome soluja, ei ole havaittu Arabidopsis lehtiä GCs (Melaragno et al., 1993), joka takaisi kloroplastilukutietojen tulkinnat solujen 2C-tasolla., Yhdessä nämä raportit viittaavat siihen, että lehtiä GCs ovat potentiaalisesti erinomainen malli systemaattinen analyysi kloroplastiin numero dynamics erityisesti solujen lineage.
Arabidopsis Leaf Guard-Solujen mallina Opiskelun Ohjaus Kloroplastiin Numero
historian GC kloroplastiin tutkimus, kloroplastiin laskenta klo ilmarakojen (GC pari) taso on palvellut yhtä tärkeä rooli määritettäessä kloroplastiin numero laskenta yksittäisten GC tasolla. Molemmat menetelmät tuottavat saman keskimääräisen klooriplastiluvun (voirasva, 1973)., Kun vaihtelu kloroplastiin jakelu pariksi GCs ja sen taustalla oleva mekanismi on aihe, tarkennus, yksityiskohtaisia tietoja viherhiukkasia yksittäisen solun tasolla, eli niiden koko, muoto, ja solunsisäisen lokalisointi on välttämätöntä. Kloroplastiin (plastid) leviämisen aikana GMC–GC eriyttäminen oli aiemmin tutkittu useita kasvilajeja (Butterfass, 1973, Butterfass 1979)., Nämä tutkimukset ehdotti kaksi mallia määrittämiseksi terminaali kloroplastiin numero GCs eri kasvilajeja (Kuva 1B): yksi (malli 1; sokerijuurikas) koskee vain kloroplastiin osiointi klo GMC jako, ja toinen liittyy paitsi kloroplastiin osiointi mutta myös kloroplastiin leviämisen aikana GC kehitystä.
aikakaudella molekyyli genetiikka, genomiikka, solujen kuvantamisen, ja muut tieteidenvälistä analyysit, on monia mahdollisuuksia edelleen luonnehdinta kloroplastiin osiointi mekanismi., Arabidopsis leaf GCs voi olla yksi parhaista mallijärjestelmistä tähän tarkoitukseen. Useat tutkimukset ovat tutkineet GC kloroplastiin numero lehtiä tai sirkkalehtien ja Arabidopsis (Hoffmann, 1968; Piikkiin ja Iilimato, 1994; Piikkiin et al., 1994; Robertson ym., 1995; Keech et al., 2007; Chen et al., 2009; Yu et al., 2009; Higaki et al., 2012; Fujiwara ym., 2018). Nämä GCs-näyttely vaatimaton määrä viherhiukkasia, jotka vaihtelevat 3,5-5,5 keskimäärin. Tähän mennessä ei ole tutkittu kloroplastin (plastid) määrän muutoksia stomataalisen kehityksen aikana., Mahataalisten kehitysanalyysien mikroskooppiset todisteet (esim. Zhao ja Sack, 1999; Hachez et al. 2011) ja alustavat havainnot osoittavat, että GMCs voi olla pienempi määrä kehittää viherhiukkasia kuin GCs ja että kloroplastiin leviämisen aikana voi esiintyä GC eriyttäminen. Testata tätä, muodostumista kloroplastiin jako koneiden GCs oli seurata koetin FtsZ1 sulatetun vihreä fluoresoiva proteiini (FtsZ1-GFP) (Fujiwara et al., 2008)., Epifluoresenssimikroskopialla (Fujiwara et al., 2017). Laajenevia lehtiruoteja (4 viikon ikäisten taimien viidennet lehdet) käytettiin. Tämän seurauksena havaittiin GCS, jossa oli symmetrisesti kuristavia klooriplasteja (Kuva 1c). Näitä viherhiukkasia muodostunut FtsZ1 rengas, kloroplastiin jako rengas strooman pinta sisempi kirjekuori kalvo lehtien mesophyll ja jalkakäytävä solut (Vitha et al., 2001; Fujiwara ym.,, 2008), Päiväntasaajan kuristusalueella. Yhdenmukainen Arabidopsis leaf development (Donnelly et al., 1999; Andriankaja ym., 2012), jakamalla kloroplasteja havaittiin myöhään, sekä varhaisessa vaiheessa, vaiheessa lehtien laajeneminen. Näin ollen malli 2 sopii mitä todennäköisimmin parhaiten Arabidopsis leaf GCs: ään.
Suhde Kloroplastiin Leviämisen Ja Laajennus Arabidopsis Lehtiä Vartija Solut,
Lisäksi, odottamaton fenotyyppi GC kloroplastiin morfogeneesin parissa oli havaittu kypsä GCs (Kuvat 1D, E)., Kun ihon kuorii täysin laajennettu lehdet (kolmas–neljäs lehtiä terät 4-viikkoa vanhoja taimia) alkaen TP-sulatettu keltainen fluoresoiva proteiini (YFP) linja olivat mikroskooppisen ominaista (FL6-5 rivi; Fujiwara et al., 2018), jotkut ilmaraot osoitti epätasainen kloroplastiin jakelu malleja GC-pareja, kun taas useimmat lehtien ilmaraot osoitti yhtä suuri tai vastaavia kloroplastiin jakauma (Robertson et al., 1995; Fujiwara ym., 2018)., Sisällä GC pari avanne, koko viherhiukkasia GC sisältää pienempi määrä viherhiukkasia oli suurempi kuin muissa GCs pari sisältävät suurempi määrä viherhiukkasia (Kuvat 1D, E). Näin GCs todennäköisesti ylläpitää solukohtaista kloroplastitilavuutta solukohtaisella tasolla solukasvun aikana. Suurentuneet klooriplastit edustivat terminaalista fenotyyppiä eivätkä enää voineet jakautua levennettyihin lehtiin. Nämä tulokset vahvistettiin vuonna useita riippumattomia kokeita, riippumatta siitä, ilmaus TP-sulatettu fluoresoiva proteiini stroomaan merkintöjä (Kuva 1F).,
Tämä GC kloroplastiin fenotyyppi on tulkita siten, että korvausjärjestelmä on kloroplastiin laajennus, joka oli hyvin dokumentoitu lehtien mesophyll solujen puutteellinen valvonta kloroplastiin jako (Pyke ja Iilimato, 1994; Piikkiin et al., 1994). Tähän mennessä vain yksi tutkimus (Ellis ja Iilimato, 1985) on todettu negatiivinen korrelaatio kloroplastiin numero ja kloroplastiin koko lehtiä mesophyll solujen vehnä, ottaa huomioon, että monissa tutkimuksissa on todettu positiivinen korrelaatio solujen määrä ja kloroplastiin numero normaali lehtiä mesophyll solut (Leech ja Piikkiin, 1988; Piikkiin, 1997)., Kun taas GC-klooriplastiluvun epätasapainoa esiintyy harvemmin (Fujiwara ym., 2018), kloroplastiin eroja GC-pareja osoittaa, että epätasa-arvoinen kloroplastiin osiointi voi laukaista ero kloroplastiin kasvun välillä villin tyypin lehtiä solujen Arabidopsis huolimatta symmetrinen solujen jakautumista.
klooriplastikorvausvaikutus GCs: ssä voi olla lievempi kuin lehtimesofyllisoluissa., GCs saattaa pystyä kestämään niukkuus tai täydellinen menetys yhteensä kloroplastiin määrä per solu vakavasti heikentynyt kloroplastiin jako mutantteja, kuten Arabidopsis arc6 ja atminE1 ja tomaatti suffulta, ottaa huomioon, että monet mutantti GCs oli vähentynyt kloroplastiin numero ja laajentuneen kloroplastiin koko samoin lehtien mesophyll solut (Robertson et al., 1995; Forth and Pyke, 2006; Chen et al., 2009; Fujiwara ym., 2018)., Myöhään kloroplastiin jako mutantti, arc5, vähennys GC kloroplastiin määrä ei liittynyt merkittävää kasvua kloroplastiin koko, toisin kuin lehtien mesophyll solut (Pyke ja Iilimato, 1994). Matalampi kloroplastiin laajennus GCs kuin mesophyll solut (Pyke ja Iilimato, 1994; Barton et al., 2016), ja vaihtelu kloroplastiin laajennus keskuudessa GCs, saattaa taustalla niin laaja sallitun alueen koko kloroplastiin määrä per GC., Lisäksi ajoitus kloroplastiin jako aikana GMC–GC eriyttäminen saattaa vaikuttaa merkittävästi terminaalin GC kloroplastiin fenotyyppi. Vaikka edelleen yksityiskohtainen luonnehdinta on tarpeen käsitellä tätä kysymystä, se näyttää uskottavalta, että Arabidopsis lehtiä GCs edustavat järjestelmän tutkimaan hyödyntämättä näkökohtia kloroplastiin numero hallita kasvien soluissa.,
toimiva Malli Kloroplastiin Määrä Päättäväisyyttä Arabidopsis Lehtiä Vartija Solut,
perusteella edellä, voimme ehdottaa toimiva malli (laajennettu malli 2) analyysi kloroplastiin numero GCs (Kuva 1G). Lopullinen kloroplastiin määrä per GC määräytyy kloroplastiin osiointi klo GMC jako ja kloroplastiin leviämisen GCs. GMC-jakautumisen aikana klooriplastit voivat olla joko samanarvoisia tai epätasa-arvoisia. Aikana kloroplastiin leviämisen, GC kloroplasteissa on lisääntyä joko yhtä (sininen viiva) tai valikoiva (magenta viiva) jako pätevyys., Esimerkiksi, jos yhtä osioitu viherhiukkasia hallussaan vastaava jako osaamisen, tasa-kloroplastiin numerot esiintyy GC pari (Kohtalo 1). Jos epätasaisesti osioitu viherhiukkasia hallussaan vastaava jako osaamisen, viherhiukkasia kasvaa samaan tahtiin sisällä GC pari (Kohtalo 3). Jos valikoiva kloroplastiin jako tapahtuu GCs, tasapaino kloroplastiin numero GC pari muuttuu, kun GMC jako (Fates 2 ja 4)., Se on tällä hetkellä vaikea löytää tukea ”valikoiva kloroplastiin jako,” mutta jos Fates 1 ja 4 itse asiassa enemmistönä GCs, niin he saattavat hallussaan mekanismi, joka ohjaa koko kloroplastiin määrä per solu, kuten lehtiä mesophyll soluja. Malli herättää kaksi kysymystä: i) onko GC-klooriplastit asianmukaisesti jaettu tytärsoluihin ja miten ne jakaantuvat? Ja ii) säännelläänkö GC-klooriplastien jakovalmiutta koordinoidusti?,
Osalta kysymys (i), onko kloroplastiin perintö tapahtuu satunnainen jakelu useita viherhiukkasia sytoplasmassa tai positiivinen kloroplastiin osiointi mekanismi(t) on ollut jo pitkään huolta (Butterfass, 1969; Birky, 1983; Hennis ja Birky, 1984; Nebenführ, 2007; Sheahan ym., 2016). Kiehtovan, Arabidopsis arc6, lehti-tai sirkkalehti GCs on nolla kolme viherhiukkasia, ja kloroplastiin-puutteellinen GCs, ei-fotosynteesin plastideissa edelleen olemassa vesicular pitkänomainen muotoja (Robertson et al., 1995; Chen ym., 2009; Fujiwara ym., 2018)., Ei GCs vailla plastideissa sinänsä on löydetty arc6, ja mitään selitystä ei annettu, vaikka häiriöitä kloroplastiin jako-laitteet (Vitha et al., 2003). Näin ollen on tärkeää tutkia, replikointi ja morfologia arc6 viherhiukkasia vuonna ilmarakojen linjaa tutkimukset. Arabidopsis-mutanttitutkimus voi antaa myös toisen vihjeen asiasta., Havainto, että 18% sirkkalehden GCs rypistynyt lehtiä (crl) mutant sisällä plastidic rakenteiden sytoplasmaan, kun taas 100% lehtiä mesophyll solut sisältävät yhdestä neljään laajentunut viherhiukkasia (Asano et al., 2004; Chen et al., 2009), on erittäin tärkeä. CRL on kloroplastinen ulompi kuoriproteiini, jonka toiminta on tuntematon. Ymmärrystä CRL-proteiinin toiminta voi tarjota oivalluksia mekanismi(t) kloroplastiin osiointi., Analyysi kloroplastiin leviämisen ja osiointi lehtiä mesophyll solujen Arabidopsis arc mutantteja ja muita siirtogeenisiä linjoja on edistänyt tutkimusta leviämisen estämistä ja eristämistä ei-mesophyll plastideissa. Samoin GCs: ssä saadut tulokset voivat olla siirrettävissä muihin solujärjestelmiin.
Huomautus
GC-malli avaa monia mahdollisuuksia kehittämiseen kloroplastiin biologia. Esimerkiksi, vaikka sytoskeletaalijärjestelmien tiedetään säätelevän kloroplastista morfologiaa, liikettä ja osiointia (Sheahan et al.,, 2016; Wada, 2016; Erickson ja Schattat, 2018), rooli kunkin sääntely-geenin kloroplastiin leviämisen ja osiointi kasveja on saanut vähän huomiota. Toisaalta, kun se on mahdollista heikentää GC kloroplastiin numero tai morfologia kautta erilaisia kokeellisia strategioita, uusia oivalluksia molekyyli valvonta kloroplastiin morfogeneesin parissa vuonna ilmarakojen lineage solujen voi olla edellyttäen. Lisäksi, yhdessä määrällisiä analyyseja kloroplastiin numero aikana ilmarakojen kehitys, matemaattinen mallinnus voi tarjota uuden avenue näihin tutkimuksiin., Tässä kirjassa esitetään nykyinen tieto siitä, miten GC kloroplastiin määrä on hallinnassa, ja korostaa, että mahdollinen hyöty on Arabidopsis lehtiä GCs ymmärrystä kloroplastiin leviämisen estämistä ja eristämistä.
tietojen Saatavuusilmoitus
Kaikki tämän tutkimuksen aineistot sisältyvät artikkeliin / lisäaineistoon.
Kirjailijasuoritukset
MF ideoi tutkimuksen ja kirjoitti käsikirjoituksen. MF ja kuten suorittaa kokeet. AS ja RI tekivät analyysit. Kaikki kirjoittajat lukivat ja hyväksyivät lopullisen käsikirjoituksen.,
Rahoitus
Tämän työn tukivat opetusministeriö, Kulttuuri -, Tiede-ja teknologiaministeri Japanissa alle KAKENHI (grant nro. 19K05831 MF ja 18K06314 RI).
eturistiriita
kirjoittajat ilmoittavat, että tutkimus on tehty ilman mitään kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdollisia eturistiriitoja.
tunnustukset
kirjoittajat kiittävät tohtori Nobuyuki Kanzawaa (Sofian yliopisto) neuvoista teoksessa.
Birky, C. W., Jr. (1983)., Sytoplasmaisten organellien osittaminen solunjakautumisessa. Int. Rev. Cytol. 15, 49-86.
Google Scholar
voita vaihtuvuus, T. (1969). Plastid jakelu mitoosin sulkeminen solun äiti solujen haploidi swedenklee (Trifolium hybridum L.). Planta 84, 230-234. doi: 10.1007/BF00388108
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar
voita vaihtuvuus, T. (1973). Plastidin jakautumisen valvonta ydin-DNA: n määrän avulla. Protoplasma 76, 167-195. doi: 10.,1007/BF01280696
CrossRef Full Text | Google Scholar
Qin, X., Rotino, G. L. (1995). Chloroplast number in guard cells as ploidy indicator of in vitro-grown androgenic pepper plantlets. Plant Cell Tissue Organ Cult. 41, 145–149. doi: 10.1007/BF00051583
CrossRef Full Text | Google Scholar
Sachs, J. (1875). Textbook of Botany. Oxford: Clarendon Press.
Google Scholar
Thomas, M. R., Rose, R. J. (1983)., Plastid-numero ja plastid-rakennemuutokset, jotka liittyvät tupakkamesofyllin protoplastiviljelyyn ja kasvien uudistamiseen. Planta 158, 329-338. doi: 10.1007/BF00397335
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar
Tirlapur, U. K., König, K. (2001). Femtosecond lähi-infrapuna laserit as a novel tool for non-invasiivisia real-time high-resolution time-lapse-kuvantaminen kloroplastiin jako elää nippu tuppi soluja Arabidopsis. Planta 214, 1-10. doi: 10.,1007/s004250100597
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar
Villi, A., Susi, G. (1980). Vaikutus eri valon voimakkuudet taajuudella ja koko ilmaraot, kokoa solujen, lukumäärä, koko ja klorofylli sisältöä viherhiukkasia vuonna mesophyll ja vartija solujen aikana ontogeny ensisijainen lehdet Sinapis alba. Z. Pflanzenphysiol. Bd. 97, 325–342. doi: 10.1016/S0044-328X(80)80006-7
CrossRef Koko Teksti | Google Scholar
















Vastaa